Zusammenfassung
Im Sommer 2022 identifizierte ein Forschungsteam der University of Wyoming eine bedeutende Kochstelle in einer spätpleistozänen Ablagerung der Bachner-Site in Zentralalaska. Dieses Sediment, das mit der Radiokohlenstoffmethode auf ca. 13.100 cal BP datiert wurde, enthält gut erhaltene Fauna von verschiedenen Klein- und Großwildarten sowie Fettsäuren, die zu hochauflösenden Ernährungsrekonstruktionen der ersten Menschen Alaskas verwendet werden können. Die Fundstelle F22-9 enthält feinkörnige kryptokristalline Silikate, die auf eine Wärmebehandlung hindeuten, identifizierbare Pflanzenreste sowie durch Feuer gebrochene Schiefer- und Quarzstücke. Die Untersuchungen des 2022 ausgegrabenen Teils der Kochstelle deuten darauf hin, dass es sich um eine der frühesten Kochstellen auf heißem Stein in Nord- und Südamerika handelt. Außerdem wurden molekulare Beweise für Fischfang geliefert, der in der Region, wenn nicht sogar auf dem gesamten Kontinent, am frühesten dokumentiert ist. Dies hat Auswirkungen auf den Lebensunterhalt, die Mobilität und die soziale Organisation der ersten Menschen Alaskas, was wiederum Aufschluss über die Ursprünge und Kolonisierungsrouten der ersten Amerikaner geben könnte. Darüber hinaus hat die frühe Bedeutung von Fischen, auf die dieser Fund hindeutet, das Potenzial, ökologische Modelle und zeitgenössische Landmanagementstrategien zu verfeinern. Zukünftige Forschungen an diesem Ort werden auf diesen ersten Schlussfolgerungen aufbauen, um zusätzliche Herdmaterialien, Mikromorphologie und Subsistenzinterpretationen auf der Grundlage traditionellen Wissens zu liefern.
Did the first Americans fish?
The question of what the first Americans ate and the technologies that they developed to meet their subsistence pursuits has been a significant topic of scholarship in North America since the discovery of the Folsom cultural tradition. After a projectile point was found embedded in a long-extinct species of bison, archaeologists have primarily conceived of the first Americans as large game specialists (Meltzer 2000; Pitblado 2021; Waguespack 2012). Some have gone even further, arguing that the first Americans caused a megafaunal blitzkrieg that led to the disappearance of most charismatic megafauna from the Americas (Martin 1984; Surovell et al. 2016).
The current picture of early American culture may be a result of research bias, however, as screen size, identification, and researcher interests may all drive an over-representation of large game hunting in the archaeological record (Lyman 1984; Partlow 2006; Speth 2010; Waguespack 2005). Investigating subsistence using a combination of traditional and novel molecular methods informed by local traditional knowledge may help to better understand the diversity of early American diets (Gifford-Gonzalez 2018). In turn, this could help to refine our interpretations of early American subsistence and cultural traditions that may contribute to our understanding of the timing and routes of initial colonization. If fishing was more important to the first Americans than is currently understood, it could bolster arguments for a coastal origin and/or coastal migration route through the continent (Braje et al. 2017; Erlandson and Braje 2022).
Fishing is a significant development in hunter-gatherer subsistence that conveys many behavioral and technological innovations (Brinkhuizen and Clason 1986). Homo habilis and Homo erectus were the first hominins to fish according to evidence from Olduvai Gorge, indicating the great antiquity of this subsistence tradition (Stewart 1994). In many regions, fishing and shellfishing are associated with sophisticated technologies and extensive knowledge of local resources, but fishing can simply represent opportunistic use of abundant resources (Erlandson 2001). Regardless, humans have pursued a wide range of aquatic resources with increasing intensity and specialization since the Upper Paleolithic and certainly by 15,000 years ago across Europe and Asia (Conard et al. 2013; Morisaki et al. 2019; Straus 1990; Zhilin 2014). This tradition certainly coincides with the arrival of the first Americans and indicates that fishing and aquatic resources may have been more important to them than current archaeological models allow.
Central Alaska, as the most likely entryway to the Americas based on decades of investigations of genetic and material evidence (Meltzer 2021; Potter et al. 2017; Raff and Bolnick 2014), is an ideal place to consider early subsistence traditions and use of fish. This region represents the nucleus of American cultures, which likely gave rise to the first Americans’ behavior, organization, and subsistence strategies. Considering subsistence traditions across the earliest occupations in this region can show the strategies that first Americans may have brought with them as they spread across the continent.
Evidence for fishing in central Alaska
The vast region between the Brooks and Alaska ranges that once extended across the Bering Strait to Siberia contains approximately 15,000 years of evidence pertaining to subsistence. Deposits in this region are known for their stratigraphic integrity and excellent preservation due to relatively cold and dry climatic conditions. Deeply stratified loess preserves the earliest cultural features and components better than any other region in the Americas (Surovell et al. 2022). These conditions provide an excellent record of resource procurement and processing or hunting and cooking traditions.
From 15,000 years ago until the arrival of the first Euro-Americans to the region approximately 150 years ago, central Alaskans have maintained a highly mobile hunting and gathering strategy (Potter 2016; Smith 2022). This allows archaeologists to draw not only from the archaeological record but the ethnographic record to make inferences about past subsistence. Linguistic and toponym data can also be brought to bear on archaeological questions (Kari 2010). The connections between the recent and deep past are valuable to our understanding of the region’s history and the history of the first Americans.
In central Alaska, the earliest sites in the region tend to be ephemeral, open-air camps that contain evidence for a wide array of food resources, including large game, waterfowl, and potentially fish (Goebel and Potter 2016; Holmes 2001; Yesner 1996). The first definitive evidence for salmon fishing is radiocarbon dated ca. 11,500 cal BP, at the site of Xaasaa Na’ (Halffman et al. 2015). This evidence is represented by burnt salmonid vertebrae as well as compound-specific isotopic evidence for fats attributable to anadromous, or migratory, salmonids that originated from the ocean (Choy et al. 2016). A paired methodology of traditional and molecular faunal analysis established through the analysis presented here might help to identify fishing during the late Pleistocene among the first Alaskans.
The importance of freshwater and anadromous (or migratory) fish resources to central Alaskans is well-documented in the present and recent past (Andrews 1988; Guedon 1971; Mishler 1986; Shinkwin 1979). Indeed, the recent shortfall of these subsistence resources has led tostate of emergency in many remote village communities in Alaska (Carothers et al. 2021), emphasizing the need to improve our understanding of the traditional, long-term relationship between various fish and people of the region. Dene speakers have likely lived in the region for the last 10,000 years and provide a continuous record of culture and traditional knowledge that can be used to interpret the archaeological record (Kari 2010; Smith 2022). Unfortunately, the regional archaeological record of fishing is largely lacking aside from the extensive evidence for fishing recovered from two early Holocene occupations at Xaasaa Na’ and potentially physical remains from late Pleistocene components Broken Mammoth that have yet to be fully analyzed (Erlandson 2001; Halffman et al. 2020). Currently, archaeologists are faced with a discontinuous record of fishing that only hints at the ultimate importance of this tradition throughout the history of human occupation in the region, from its origins to potential connections with the present.
While archaeologists have successfully identified small game and avifauna at many of the earliest sites in central Alaska (Goebel and Potter 2016), definitive evidence of fish has yet to be recovered in faunal assemblages dating to the late Pleistocene. This may be due in part to bias present in the deposition, preservation, and analysis of assemblages. In central Alaska in particular, bone recovered at sites from all cultural periods tends to be highly fragmentary, likely related to processes of cooking or cleaning that continued into the period following Euro-American contact (Potter 2007). This results in assemblages where only a small fraction of the total faunal material is identifiable to general taxonomic class, and the remains that are identifiable tend to be from larger game (Grayson 1984; Lyman 2019). Additionally, the cleaning of fish traditionally took place in bodies of water, where skeletal remains could be directly discarded in rivers or lakes before the cleaned fillets were transported to another location for cooking (Nelson 1983).
In the rare cases where identifiable fish remains were deposited, researchers may not have recovered or identified them. Alaskan and North American researchers have not consistently used screens or mesh small enough to capture the remains of smaller fauna, so some assemblages may not contain these small remains (Cook 1977; Holloway et al. 2018; Shaffer 1992). This practice would result in few deposited fish remains at residential sites. Research in the region has primarily focused on the analysis of stone tool technology rather than detailed faunal analyses (Goebel and Potter 2016). It is well-documented that technologies used in the harvest and preparation of fish in Alaska and elsewhere are largely organic, not lithic (Erlandson 2001; Osgood 1937). Zooarchaeological training and reference collections are focused on mammalian identification at many institutions (Betts et al. 2011), leaving North American analysts ill-prepared for fish identification (Lau and Kansa 2018; Peres 2010). This mammal-centric view of subsistence may contribute to a biased picture of resource procurement in central Alaska from the late Pleistocene to the present.
Applying molecular faunal identification methods to other, earlier sites in the region can help to connect the dots between what we know from the most recent periods in the region’s history, the early Holocene, and the first Americans. Molecular methods can overcome the limited visibility of fish remains in faunal assemblages from late Pleistocene sites in central Alaska because they target residues from substrates like lipids, which are produced during food processing and preserve very well in central Alaska’s subarctic environment (Buonasera et al. 2019; Kedrowski et al. 2009; Taché and Craig 2015). The compound-specific isotopic analysis of specific fats, palmitic acid (C16:0) and stearic acid (C18:0), can be used to distinguish between common local taxa to reconstruct past diets (Buonasera et al. 2015; Choy et al. 2016). These fats can be deposited on cooking surfaces like pottery or in sediments related to hearth cooking features and can be extracted using solvents (Buonasera et al. 2019; Craig et al. 2007). Esterified fatty acids or fatty acid methyl esters (FAMEs) are then submitted for isotopic analysis of carbon, which can be used to discriminate between terrestrial, lacustrine, and marine/anadromous dietary sources (Choy et al. 2016; Taché and Craig 2015). Therefore, an analysis of sediment FAMEs can reveal dietary patterns in situations where bones may not be identifiable, may not have been deposited at the site, or may not have been collected under certain excavation protocols.
Finding fishing through traditional and molecular Methods
Through a mixed methods approach to dietary reconstruction, researchers can combine data from physical faunal remains to understand expected animal contributions to diet and pair them with molecular data that can reconstruct contributions of different dietary sources. Identifying physical faunal remains is a traditional approach to zooarchaeology that has been employed in the field for decades by both North American- and European-trained researchers (Gifford-Gonzalez 2018). Research institutions around the world prioritize specialists who can identify animal bones in hiring because it has long been recognized as an essential analytical contribution to the field (Trigger 2006). Many renowned institutions have reference collections of hundreds or thousands of animal skeletons to facilitate this comparative analysis. Despite the limitations of this analysis in central Alaska, it is still a very useful tool for ascertaining diet breadth at different periods in this region.
Molecular methods have become more important in dietary reconstruction and zooarchaeology in the last two decades (Steele 2015). Isotopic dietary reconstruction is now commonplace in studies of human remains and is becoming more common as a tool for environmental reconstruction when applied to animal remains at archaeological sites (DeNiro and Epstein 1976; Halffman et al. 2020; Hoppe 2006; Schoeninger 1989). In recent years, molecular archaeologists have applied proteomic methods to identify bones and this technique, zoology by mass spectrometry or ZooMS, becomes more efficient and accurate every year (Buckley et al. 2009; Richter et al. 2022). Analysis of FAMEs via compound-specific isotopic analysis and gas chromatography/mass spectrometry follows these trends as a way to indirectly reconstruct faunal assemblages through molecular components preserved at archaeological sites (Meier-Augenstein 2002).
Combining traditional and molecular analysis leverages the strengths of both methods. Traditional methods are excellent for understanding the animals processed on-site. Molecular methods can employ the results of this analysis to show how much of each resource was used (Buonasera et al. 2015; Taché and Craig 2015). Zooarchaeologists have long debated the correspondence between aggregate measures like minimum number of individuals (MNI) and number of identifiable specimens (NISP) and diet (Grayson 1984; Lyman 2019; Marshall and Pilgram 1993; Morin et al. 2016). Molecular methods like the compound-specific isotopic analysis of FAMEs can overcome these limitations through Bayesian mixing models that show the expected contribution of different sources indicated by traditional analysis to the resulting isotopic signature (Choy et al. 2016). Researchers can easily compare these results across sites to elucidate regional dietary patterns.
Bachner Site, Theech’el Menn’: Window into central Alaskan cooking
The Bachner Site (XBD-155) provides an excellent opportunity to apply this mixed-methods approach because it offers multiple components spanning the late Pleistocene to the historic period, relatively well-preserved faunal remains, and several large cooking features with well-preserved fatty acids. This site, located since at least the early Holocene on a large lake, is situated in the middle Tanana Valley and just south of the Shaw Creek Flats, both offering salmon-bearing streams less than 2 km away (Fig. 1). It sits on a rise approximately 50 m above the lake today and likely overlooked a wetland during the late Pleistocene. Neighboring sites include Swan Point (15 km), Broken Mammoth (16 km), Mead (15.5 km), and Holzman (15.5 km), each of which exhibits late Pleistocene cultural occupations that date to the earliest period of American history (Gilbert 2011; Holloway 2016; Lanoë and Holmes 2016; Wygal et al. 2018). These are all open-air sites; cave occupations are very rare in this region (Hoffecker and Frederick 1996).

The Bachner Site was first identified by Charles Holmes and Richard VanderHoek in 1990, who chose to visit the site based on its prominence on the north shore of the lake (Gelvin-Reymiller and Reuther 2010). Through surface collection and a 1 x 1 m test unit, they established that the site had multiple components in well-stratified loess. Carol Gelvin-Reymiller conducted additional testing at the site, including six shovel tests and a 1 x 2 m unit that expanded on the original test unit. These results were incorporated in Reuther’s (2013) dissertation research and showed that the site had at least four distinct cultural occupations spanning the late Pleistocene to the present (Fig. 2).

In 2021–2022, a team from the University of Wyoming undertook a comprehensive GPR and auger survey at the site to identify areas most likely to yield cooking features (Doering et al. in review). Three areas were targeted based on shifts in dielectric constant and high projected artifact density (Fig. 3). Excavations were carried out in 5 cm levels by 50 x 50 cm quad in 1 m² units. Features, diagnostic or large (> 5 cm) bones, and diagnostic or large (> 2 cm) lithic material were collected in situ and piece-plotted using a Geomax Zoom 10 Total Station. All other material was screened through ⅛th inch hardware cloth and collected according to type, level, and quad. Materials were returned to the University of Wyoming for cataloging, curation, and analysis.

Analysis of hearth sediments
Sediments from hearth or cooking features were identified in the field by the presence of oxidized or burnt sediment, ash, charcoal flecking, and/or burnt artifacts (Black and Thoms 2014). Feature sediments related to cooking were piece-plotted, photographed, and collected in bulk by quad and level. Bulked sediments from were screened through 150 μm geologic screens and material > 150 μm was reserved for faunal, lithic, and paleobotanical analysis. Paleobotanical specimens were analyzed by Katherine Puseman (PaleoWest) through a comparison to representative regional samples to identify hearth fuels and possible cooked plant material (Carlquist 2013; Delorit 1970; Hoadley 1990; Martin and Barkley 1961; Panshin and Zeeuw 1970). Puseman examined material under a Bausch and Lomb Stereozoom microscope at magnifications of 10-70x.
Fatty acid methyl esters were extracted from sediments with dichloromethane using a modified Bligh-Dyer protocol (Buonasera et al. 2015; Choy et al. 2016; Doering et al. 2020; Kedrowski et al. 2009). For each sample, 2 g of hearth sediment was weighed out into a clean conical 15 mL glass centrifuge tube and 5 μg internal standard was added (1000ppm nonadecanoate). Lipids were extracted from sampled sediment with 10 mL DCM:M (dichloromethane:methanol; v/v 1:1) through sonication (20 minutes) and centrifugation (15 minutes at 4,000 RPM). The supernatant was filtered through glass wool into a clean glass centrifuge tube. This process was repeated once, and the resulting supernatant was combined into one centrifuge tube which was centrifuged once more and filtered into 50 mL glass scintillation vials. These were placed under a gentle stream of nitrogen until all solvent had evaporated and solvents were dried. The total lipid extract was weighed so that yields could be compared to expected values.
Methyl esters were derivatized by adding 5 mL sulfuric acid (4 %) in methanol and heating to 85 °C for 60 minutes. After cooling for 20 minutes, 5 mL sodium bicarbonate solution was added to quench the reaction, and derivatized lipids were extracted with 3 mL of hexanes. Samples were then vortexed, and the supernatant was removed to a new 15 mL glass sample tube. Hexane addition, vortexing, and supernatant removal were repeated two more times. Vials containing derivatized lipids were placed on a heat block set to 30 °C and under a gentle stream of nitrogen for 30 minutes. Dried FAMEs were weighed, and 1 mL of dichloromethane was added to each. The diluted FAMEs were transferred to a new gas chromatography-mass spectrometry (GC/MS) autosampler vial and analyzed at the University of Wyoming on a Trace 1310-ISQ with a gas chromatographer with a DB5 column to confirm the presence of internal standard, C16:0, and C18:0. Dried aliquots to the University of California, Davis for compound-specific isotopic analysis via gas chromatography-combustion-isotopic ratio mass spectrometry.
Compounds were analyzed on a Trace 1310 gas chromatograph coupled to a Thermo MAT 253 isotopic ration mass spectrometer through a GC IsoLink II combustion interface. Samples were injected, splitless, on a DB-5 ms column (60 m x 0.25 mm OD, 0.5 mm film thickness; constant flow 1.4 mL/min). Once separated, FAMEs were quantitatively converted to CO2 in with a NiO/CuO catalyst at 1000 °C, dried, and introduced to the isotopic ratio mass spectrometer. Provisional isotopic ratio mass spectrometer values were corrected both based on working standards composed of FAMEs calibrated against NIST standard reference materials, and for the isotopic contribution of methanol, with a resulting standard deviation of ±0.11 ‰ or better. International standards are set for carbon and nitrogen isotopes: Vienna Pee Dee Belemnite ˇ(VPDB) and AIR, respectively. These standards represent the ratio of 13C to 12C and 15N to 14N, and this ratio is noted in delta units per mil (δ13C ‰ and δ15N ‰).
Previous research has shown that the isotopic values of N, C16:0, and C18:0 in terrestrial and aquatic fauna are significantly different and suitable for stable isotope mixing models (Choy et al. 2016). Isotopic contributions of various fauna were estimated using δ13C isotopic values of C16:0 and C18:0 using background compound-specific isotope data collected from subarctic fauna and corrected post-industrial carbon (Taché and Craig 2015; Choy et al. 2016; Buonasera et al. 2015). An open-source package in R, simmr version 0.5.0.9000, was used to estimate relative contributions of potential dietary contributions (Parnell and Govan 2023). Its Bayesian statistical framework incorporates uncertainty in modeling different food groups, making it ideal for estimating the relative contributions of dietary items in hearth remains. Following Choy et al. (2016), lacustrine resources represent values from both waterfowl and lake fish.
Dietary and environmental data from the Late Pleistocene
Three definitive hearth features were recovered during the 2022 season, including a significant ashy feature from the late Pleistocene component at the site (F22-9), approximately 165-180 cm below surface. This feature, identified in an auger test and excavated as a stand-alone 1 x 1 test unit, had burnt lithic material, fire-cracked rock related to cooking, hundreds of faunal fragments, and charred plant remains including charcoal (Fig. 4). The feature is bowl-shaped, indicating that it was excavated into the surrounding substrate, and approximately 10 cm deep at its thickest (Fig. 5). This feature was only partially excavated, and 2023 excavations expanded the area around the initial test to record the feature’s overall size and shape, revealing that it is approximately 1.5 m in overall diameter and 13 cm thick at the center.


Molecular results
Two contexts were sampled from F22-9 for molecular material. The first sample was taken from a bulk sediment sample collected from the first 5 cm of the feature excavation (FS 1901) and the second was collected from the second level, or middle 5 cm of the feature excavations (FS 2009), which contained ashier sediments and possibly reflects the hotter parts of the hearth feature (Braadbaart et al. 2012; Thoms 2008). Both samples were duplicated and run in parallel with a sample blank. Results show average δ15N = 4.8 ±0.001, δ13CC16:0 = 29.90±0.52, and δ13CC18:0 = -29.84 ± 0.20 (Table 1). Compared to known values of animal resources in the area, the values appear to represent a mix of lacustrine and terrestrial fauna (Fig. 6).

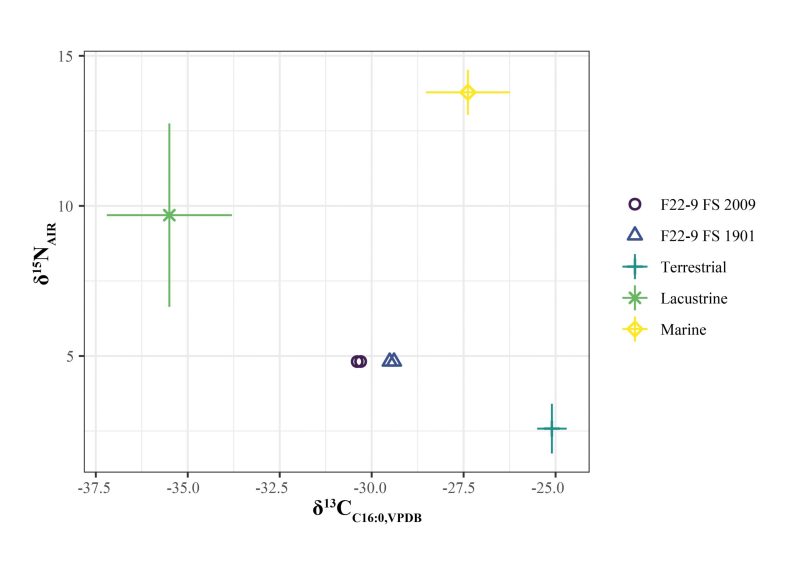
An unpaired t-test was conducted to compare the isotopic composition in the sampled sediments. The feature sediment collected from the center of the feature exhibited significantly lower δ13C values in palmitic acid (C16:0, -0.895 ‰, p < 0.001). Results of t-tests on δ15N and δ13C of stearic acid (C18:0) did not show any significant differences (p = 1.0 and p = 0.25, respectively). This suggests that lipids recovered from the center of the hearth, possibly the hottest part of the hearth, do exhibit differences in palmitic acid carbon isotopic composition but not stearic acid carbon or bulk nitrogen values.
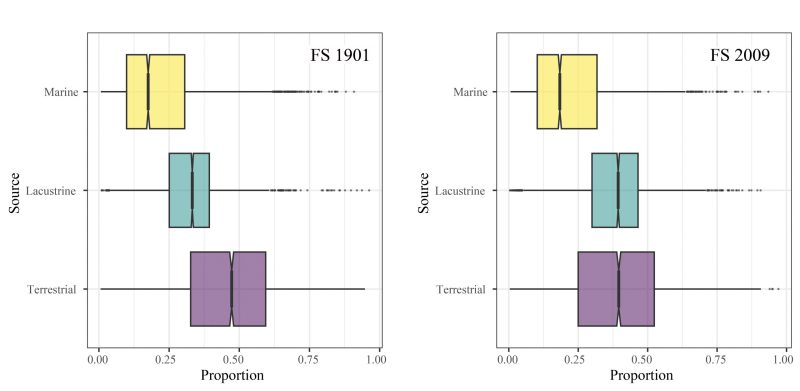
Applying a Bayesian mixing model to these data shows the relative estimated contributions of different dietary inputs to the measured bulk and compound-specific isotopic signatures from hearth sediments. These indicate that terrestrial fauna was the primary contributor to isotopic signatures, but aquatic fauna such as lacustrine fish and waterfowl also likely contributed to the fatty acids recovered from this feature (Fig. 7). Ongoing analysis of physical faunal remains indicates that Lagopus spp. (grouse), Urocitellus parryii (arctic ground squirrel), and Bison priscus (steppe bison) were cooked in the hearth. No definitive bone from fish or waterfowl has been recovered and identified in contrast to mixing model results.
Paleobotanical results
Four samples of floated light fraction were analyzed by Katherine Puseman at Paleoscapes. These were each separated into 1 mm and 0.5 mm fractions. Approximately 9.8 g of paleobotanical material was recovered in 2022 (Table 2). Salicacae (willow or cottonwood) was the most common burnt plant remain identified, comprising 22 % of the total assemblage by weight. Other possible fuels included Betula spp. (birch), but this only represented 1 % of the total assemblage by weight. Salicacae provided the bulk of the fuels for the cooking feature.

Edible plant materials were also recovered, including four charred seeds of knickiknick or bearberry (Arctostaphylos spp.), which typically fruits in the late summer, and processed edible tissue of a likely fruit or berry (Stenholm et al., 1994). One geophyte was also identified and recovered and could relate to several local edible plants with underground storage organs. These might include Eskimo potato (Hedysarum alpinum), wild onion (Allium schoenoprasum), or cattails (Typha latifolia) (Holloway 2016; Kari 1987). Finally, one seed of the Fabacaea family was identified, possibly belonging to Nootka lupine (Lupinus nootkatensis), which has edible roots (Holloway 2016: 185). These results indicate that plants were also cooked in F22-9.
What was cooking during the late Pleistocene?
Research at the Bachner Site is ongoing, and analysis of materials will be incomplete until the late Pleistocene feature (F22-9) is fully excavated. However, several important trends are already apparent in the molecular and paleobotanical data presented above. Macrobotanical and zooarchaeological evidence provides a proxy for past environment at the site, suggesting that the local ecology matches broader regional trends that have been reconstructed through pollen and isotopic studies (Bigelow and Powers 2001; Kaufman et al. 2016). Woody taxa identified in the hearth feature include shrubby species associated with the drier, colder late Pleistocene subarctic environment. A fragment of an arctic ground squirrel mandible recovered in the feature along with a femoral head from a now-extinct Bison priscus also reflect a treeless, shrub tundra environment. Isotopic signatures suggest that the local environment was marshy, with waterfowl and/or fish species present in the area.
The resources preserved in F22-9 also provide evidence for subsistence and cooking techniques employed by early Alaskans. Late Pleistocene residents at the site were cooking a wide range of fauna and flora, including small game, large game, late summer berries, and starchy plants. Molecular results also strongly suggest that aquatic resources such as lake fish were cooked in this feature. The people who inhabited the site used available woody taxa, including willow or cottonwood and birch, likely dwarf birch (Betula glandulosa), to cook the food in this feature. Salicacae are soft hardwoods that do not produce as much heat as hardwoods such as spruce (Picea spp.) or paper birch (Betula neoalaskana). Nevertheless, with a heating value of 19.8 MJ/od kg, these species could have efficiently fueled a subterranean cooking feature that gradually cooked a diverse array of plant and animal taxa (Black and Thoms 2014). Willow and cottonwood are also commonly used in many fish preparation techniques in the ethnographic present (Holloway and Alexander 1990, 222), and their near ubiquity in the charcoal of F22-9 provides another line of evidence suggesting that this feature was used to prepare fish.
Reconstructing cooking techniques and implications for social organization
Experimental and ethnographic research has demonstrated that hearths and cooking features vary morphologically in ways that can be used to identify different culinary traditions (Thoms 2008). Open air cooking facilities, such as cook stone grills and spit roasting, represent expedient cooking techniques that process food over a high-temperature fire relatively quickly with relatively little material and temporal investment (Thoms 2009). Earth oven and pit features, in contrast, represent greater material and temporal investments that can be more effective in processing certain plants and animals (Black and Thoms 2014), particularly starchy plants. The greater investment required by these features is typically associated with communal activities, such as feasting, and longer occupations.
In contrast with other documented hearths in the region, this feature is notably large and contains large fragments of burnt rock unrelated to lithic production (Fig. 4c). Schist and coarse-grained volcanic material show characteristic indicators of heat treatment (Neubauer 2018), indicating that these were employed in hot rock cooking (Black and Thoms 2014). While unpublished hearth features may contain similar materials (Graf et al., 2023), F22-9 is the only feature from this period documented to date that exhibits examples of fire cracked rock that likely pertained to cooking. Moose Creek possibly contains a rock-lined feature in a cultural component radiocarbon dated to 13,100 cal BP (Pearson 1999), but excavation, recording, and collection methods at this site provide only equivocal evidence for hot rock cooking. Therefore, the Bachner Site’s F22-9 currently represents the only evidence of hot rock cookery from the late Pleistocene. In combination with the wide range of fauna and flora recovered in this large feature, this strongly suggests that this feature represents an early roasting pit that combined terrestrial, lacustrine, and vegetable resources (Carney et al. 2022, 130).
Where late Pleistocene features in central Alaska typically resemble expedient open-air cooking facilities, the feature at Bachner appears to represent a closed cooking feature used to steam or slow cook many types of food over a longer period of time (Thoms 2009). Current conceptions of the first Alaskans revolve around ephemeral hunting camps, indicating small groups who exercised a strategy of high residential mobility. Molecular and material evidence from the Bachner Site points to a greater diversity of mobility strategies, with occasional logistical mobility and group aggregations. This research shows the benefit of looking beyond fragmentary faunal remains to reconstruct past dietary and culinary patterns and may help to refine conceptions of subsistence among the first Americans.
Directions for future cooking research
Additional research at the Bachner Site can show how the overall site size, subsistence evidence, and record of exchanged raw materials compares to other late Pleistocene sites in the region. Micromorphological sampling of the feature in profile took place in 2023 and reconstruction of depositional sequences is ongoing. If the feature was utilized more than once, the micromorphology should reflect this, and it could explain differences in isotopic composition documented above. Future isotopic sampling could target different morphological levels to show how resources were used at different times during the late Pleistocene. Additionally, comparing these results to other features recovered from subsequent occupations (mid- and late Holocene) may help to illustrate broader trends in environment and diet. A diachronic and regional comparison may show that subsistence and mobility were more variable than current archaeological reconstructions indicate.
This study represents a successful integration of isotopic and traditional dietary reconstruction techniques and shows several areas where this method can be advanced. Isotopic sampling is not new in archaeology, but applications of compound-specific sampling are relatively recent. Methods and reference datasets are still being refined. Plant signatures represent one of the biggest gaps in current analyses, and these have yet to be incorporated into archaeological mixing models. Lacustrine species should be further tested to determine how to better distinguish waterfowl and fish taxa signatures. More experimentation is needed to determine how fatty acids and other residues change in different cooking conditions, which could have also led to the significant difference in the carbon isotopic composition of palmitic acids documented above (Doering 2017). Researchers are also investigating how mass spectrometry may be used to distinguish between different cooked foods (Buonasera et al. 2019; Kedrowski et al. 2009). As references are added, techniques are standardized, and experiments continue, previous results can be reinterpreted.
Future research that combines bulk and compound-specific isotopic analysis of fatty acids from hearth sediments across the region and spanning human history can improve our understanding of past diets, cooking traditions, and environmental history. Working with local Indigenous communities to understand how traditional cooking practices may appear in the archaeological record would also help archaeologists to identify and document culinary traditions in the past. Improved recording and collection strategies may help to identify a greater diversity of hot rock cooking techniques. Together, ethnographic and archaeological lines of evidence would shed light on intersections between subsistence traditions and ecology. Previous research in regions near and far has shown how similar lines of evidence can be mobilized to improve ecological management strategies (Armstrong et al. 2023; Ellis et al. 2021; Lepofsky and Armstrong 2018; Moss et al. 2016; Snitker et al. 2022). Using the same line of inquiry, sustainable plans for aquatic resources can be refined in central Alaska.
Conclusion
Returning to the original question, evidence for fishing during the late Pleistocene is still equivocal but results from the Bachner site strongly suggest that fish were part of the broad subsistence round of the first Americans. Molecular and paleoethnobotanical evidence both point to fish and fish preparation. Parallels between the hot rock cooking feature identified at the Bachner Site and similar features across the northwest coast strongly suggest a feature that was built to cook several food resources, frequently including fish. This suggests that the diet of the first Americans living in central Alaska was more varied than has previously been considered. Combining traditional and molecular results is indeed an effective way to overcome possible taphonomic bias and develop our understanding of the past.
The broader subsistence system reflected by data from the Bachner Site as well as other sites in the middle Tanana Valley strongly suggests that researchers of Clovis and possible pre-Clovis culture in North America should expand their view of dietary diversity. Rather than a narrow focus on big game, all zooarchaeological evidence suggests that the first Americans ate a broad diet that incorporated a range of fauna and flora. Moreover, the late Pleistocene cooking feature at the Bachner Site suggests the variety of cooking traditions that archaeologists should look for in association with early American subsistence pursuits. Not only were central Alaskans cooking through direct fire, but it also appears that they were intentionally constructing roasting pits that could effectively slow-cook a variety of resources simultaneously. This would have required a significant investment in time and resources and a level of social organization as yet undocumented in this region at this time.
The archaeological data presented here pertain to the distant past but do have salience for central Alaskan communities in the present. Documenting the legacy of fishing traditions in the region allows land managers to understand the antiquity of fish subsistence, which can refine ecological models. Identifying the origins of roasting pits, smoking, fermentation, and other culinary traditions now common throughout the region can show how resource use changed through time. A greater understanding of subsistence, including cooking traditions, can promote ecological and heritage management. Appreciating the wide range of preparation techniques and foods processed throughout the subarctic past will help us to preserve these important traditions for many years into the future.
Acknowledgments
I am grateful to the local descendant communities of Nenana and Healy Lake for sharing their time and traditional knowledge with me, particularly Evelynn Combs, Eva Dawn Burke, and Don Charlie. To all members of the Dene community who have shared their knowledge in stories, books, and documentaries, maasee’ choo! Bea Bachner has given three generations of archaeologists permission to excavate at the site on her property. Joshua Reuther and Scott Shirar at the University of Alaska Fairbanks Museum of the North have granted access to collections and helped with landowner permissions. Charles Holmes has shared a great deal of knowledge on this site specifically and the archaeology of the region. Two field crews of University of Wyoming generated the excavation data presented here, with special thanks to Molly Herron, Sarah Allaun, Ken Hladek, Grace Stanford, Josie Corbett, and Daniel Garner. Kathy Puseman at Paleoscapes provided incredibly professional paleobotanical analysis and conservation. Jamie McFarland runs the GC/MS lab at the University of Wyoming and provided expert consultation on the protocol employed here. Funding for this research was provided by the Frison Institute, the University of Wyoming Research Office, and the College of Arts and Sciences, with special thanks to the University of Tübingen and EiszeitQuell laut Webseite for the invitation to share this research. Finally, this paper is dedicated in loving memory to Carol Gelvin-Reymiller, who cared about this place and this research so very much.
References
Andrews, E. 1988: The harvest of fish and wildlife for subsistence by residents of Minto, Alaska. Alaska Department of Fish and Game, Subsistence Division, Fairbanks, AK.
Armstrong, C. G., Lyons, N., McAlvay, A. C., Ritchie, P. M., Lepofsky, D., and Blake, M. 2023: Historical ecology of forest garden management in L a xyuubm Ts’msyen and beyond. Ecosystems and People 19, 2160823. https://doi.org/10.1080/26395916.2022.2160823.
Betts, M. W., Maschner, H. D., Schou, C. D., Schlader, R., Holmes, J., Clement, N., and Smuin, M. 2011: Virtual zooarchaeology: Building a web-based reference collection of northern vertebrates for archaeofaunal research and education. Journal of Archaeological Science 38, 755-e1.
Bigelow, N. H. and Powers, W. R. 2001: Climate, vegetation, and archaeology 14,000–9000 cal yr BP in central Alaska. Arctic Anthropology 171–195.
Black, S. L. and Thoms, A. V. 2014: Hunter-gatherer earth ovens in the archaeological record: Fundamental concepts. American Antiquity 79, 203–226.
Braadbaart, F., Poole, I., Huisman, H. D., and van Os, B. 2012: Fuel, fire and heat: An experimental approach to highlight the potential of studying ash and char remains from archaeological contexts. Journal of Archaeological Science 39, 836–847.
Braje, T. J., Dillehay, T. D., Erlandson, J. M., Klein, R. G., and Rick, T. C. 2017: Finding the first Americans. Science 358, 592–594.
Brinkhuizen, D. C. and Clason, A. T. 1986: Fish and archaeology: Studies in osteometry, taphonomy, seasonality, and fishing methods, BAR International Series. Oxford, England: British Archaeological Reports.
Buckley, M., Collins, M., Thomas-Oates, J., and Wilson, J. C. 2009: Species identification by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry: Species identification of bone collagen using MALDI-TOF-MS. Rapid Commun. Mass Spectrom. 23, 3843–3854. https://doi.org/10.1002/rcm.4316.
Buonasera, T., Herrera-Herrera, A.V., and Mallol, C. 2019: Experimentally derived sedimentary, molecular, and isotopic characteristics of bone-fueled hearths. J Archaeol Method Theory 26, 1327–1375. https://doi.org/10.1007/s10816-019-09411-3.
Buonasera, T., Tremayne, A. H., Darwent, C. M., Eerkens, J. W., and Mason, O. K. 2015: Lipid biomarkers and compound specific δ13C analysis indicate early development of a dual-economic system for the Arctic Small Tool tradition in northern Alaska. Journal of Archaeological Science 61, 129–138.
Carlquist, S. 2013: Comparative wood anatomy: systematic, ecological, and evolutionary aspects of dicotyledon wood. Springer Science & Business Media.
Carney, M., Guedes, J. d., Wohlgemuth, E., and Tushingham, S. 2022: Bulbs and biographies, pine nuts and palimpsests: Exploring plant diversity and earth oven reuse at a Late Period plateau site. Archaeol Anthropol Sci 14, 130. https://doi.org/10.1007/s12520-022-01588-1.
Carothers, C., Black, J., Langdon, S. J., Donkersloot, R., Ringer, D., Coleman, J., Gavenus, E. R., Justin, W., Williams, M., and Christiansen, F. 2021: Indigenous peoples and salmon stewardship: a critical relationship. Ecology and Society 26, 1. https://doi.org/10.5751/ES-11972-260116.
Choy, K., Potter, B. A., McKinney, H. J., Reuther, J. D., Wang, S. W., and Wooller, M. J. 2016: Chemical profiling of ancient hearths reveals recurrent salmon use in Ice Age Beringia. Proceedings of the National Academy of Sciences 113, 9757–9762.
Conard, N. J., Kitagawa, K., Krönneck, P., Böhme, M., and Münzel, S. C. 2013: The importance of fish, fowl and small mammals in the Paleolithic diet of the Swabian Jura, Southwestern Germany. In: J. L. Clark and J. D. Speth (eds.), Zooarchaeology and modern human origins: Human hunting behavior during the Later Pleistocene. Vertebrate Paleobiology and Paleoanthropology. Springer, New York, NY, 173–190.
Cook, J. P. (ed.), 1977: Archeological investigation along the Trans-Alaska Pipeline. University of Alaska, Institute of Arctic Biology, Fairbanks, AK.
Craig, O. E., Forster, M., Andersen, S. H., Koch, E., Crombé, P., Milner, N. J., Stern, B., Bailey, G. N., and Heron, C. P. 2007: Molecular and isotopic demonstration of the processing of aquatic products in northern European prehistoric pottery. Archaeometry 49, 135–152.
Delorit, R. J. 1970: Illustrated taxonomy manual of weed seeds. Agronomy Publications.
DeNiro, M. J. and Epstein, S. 1976: You are what you eat (plus a few‰): The carbon isotope cycle in food chains. Geological Society of America 6, 834.
Doering, B. N. 2017: Measuring the potential influence of cooking on the carbon and nitrogen isotopic composition of spawning Chinook salmon. Journal of Archaeological Science: Reports 12, 491–498.
Doering, B. N., Esdale, J. A., Reuther, J. D., and Catenacci, S. D. 2020: A multiscalar consideration of the Athabascan migration. American Antiquity 85, 470–491. https://doi.org/10.1017/aaq.2020.34.
Doering, B. N., Hladek, K., Herron, M., Reuther, J. D., Esdale, J. A., Holmes, C. E., Smith, and Gerad M. in review. Evaluating systematic use of ground penetrating radar and auger surveys to determine activity areas at three open air sites in Central Alaska. Journal of Field Archaeology.
Ellis, E. C., Gauthier, N., Klein Goldewijk, K., Bliege Bird, R., Boivin, N., Díaz, S., Fuller, D. Q., Gill, J. L., Kaplan, J. O., Kingston, N., Locke, H., McMichael, C. N. H., Ranco, D., Rick, T. C., Shaw, M. R., Stephens, L., Svenning, J.-C., and Watson, J. E. M. 2021: People have shaped most of terrestrial nature for at least 12,000 years. Proc Natl Acad Sci USA 118, e2023483118. https://doi.org/10.1073/pnas.2023483118.
Erlandson, J. M. 2001: The archaeology of aquatic adaptations: Paradigms for a new millennium. Journal of Archaeological Research 9, 287–350.
Erlandson, J. M. and Braje, T. J. 2022: Boats, seafaring, and the colonization of the Americas and California Channel Islands: A response to Cassidy (2021). California Archaeology 0, 1–9. https://doi.org/10.1080/1947461X.2022.2125711.
Gelvin-Reymiller, C. and Reuther, J. D. 2010: Shaw Creek Flats Easts Project, Quartz Lake Sites XBD-00362, 00361, 00159, 00155, Second Year Report (2010), RE: Field Archaeology Permit 2009-01 (File 3420-2009) (Annual Report to the State Historic Preservation Office). University of Alaska Fairbanks, Fairbanks, AK.
Gifford-Gonzalez, D. 2018: An introduction to zooarchaeology. Springer.
Gilbert, P. J. 2011. Micromorphology, site spatial variation and patterning, and climate change at the Mead Site (XBD-071): A multi-component archaeological site in interior Alaska. University of Alaska Fairbanks.
Goebel, T. and Potter, B. A. 2016: First traces: Late Pleistocene human settlement of the Arctic. In: T. M. Friesen and O. K. Mason (eds.), The Oxford Handbook of the Prehistoric Arctic. Oxford University Press, New York, NY, 223–253.
Graf, K., Shelley, N., Esdale, J. A., and Goebel, T. 2023: Stratigraphy and radiocarbon dating at the Shég’ Xdaltth’í’ site in Central Alaska. Presented at The 88th Annual Meeting of the Society for American Archaeology. 2023 (tDAR id: 473569).
Grayson, D. K. 1984: Quantitative Zooarchaeology: Topics in the analysis of archaeological faunas. Academic Press, Orlando, FL.
Guedon, M.-F. 1971: People of Tetlin, why are you singing? A study of the social life of the Upper Tanana Indians. Ph.D. Thesis, Bryn Mawr College, Pennsylvania.
Halffman, C. M., Potter, B. A., McKinney, H. J., Finney, B. P., Rodrigues, A. T., Yang, D. Y., and Kemp, B. M. 2015: Early human use of anadromous salmon in North America at 11,500 y ago. Proceedings of the National Academy of Sciences 112, 12344–12348.
Halffman, C. M., Potter, B. A., McKinney, H. J., Tsutaya, T., Finney, B. P., Kemp, B. M., Bartelink, E. J., Wooller, M. J., Buckley, M., Clark, C. T., Johnson, J. J., Bingham, B. L., Lanoë, F. B., Sattler, R. A., and Reuther, J. D. 2020: Ancient Beringian paleodiets revealed through multiproxy stable isotope analyses. Sci. Adv. 6, eabc1968. https://doi.org/10.1126/sciadv.abc1968.
Hoadley, R. B. 1990: Identifying wood: Accurate results with simple tools. Taunton Press, Newtown, CT.
Hoffecker, J. F. and Frederick, H. W. 1996: Introduction to the archaeology of Beringia. American beginnings: The prehistory and Palaeoecology of Beringia 149–153.
Holloway, C., Baxter-McIntosh, J., Coffman, S., Shirar, S., and Reuther, J. D. 2018: Mitigation report for the Trans-Alaska Pipeline Archaeological Project (1969-1977) Phase II: Archaeological inventory. University of Alaska Fairbanks Archaeology Department, Fairbanks, AK.
Holloway, C. R. 2016: Paleoethnobotany in interior Alaska. Master’s thesis. University of Alaska Fairbanks, Alaska.
Holloway, P. S. and Alexander, G. 1990: Ethnobotany of the Fort Yukon Region, Alaska. Econ Bot 44, 214–225. https://doi.org/10.1007/BF02860487.
Holmes, C. E. 2001: Tanana River Valley archaeology circa 14,000 to 9000 BP. Arctic Anthropology 154–170.
Hoppe, K. A. 2006: Correlation between the oxygen isotope ratio of North American bison teeth and local waters: Implication for paleoclimatic reconstructions. Earth and Planetary Science Letters 244, 408–417.
Kari, J. 2010: The concept of geolinguistic conservatism in Na-Dene prehistory. In: The Dene-Yeniseian Connection: Bridging Asia and North America, New Series. University of Alaska, Department of Anthropology, Fairbanks, AK, 194–222.
Kari, P. R. 1987: Tanaina plantlore, Dena’ina K’et’una: An ethnobotany of the Dena’ina Indians of Southcentral Alaska. National Park Service, Alaska Region, Anchorage, AK.
Kaufman, D. S., Axford, Y. L., Henderson, A. C. G., McKay, N. P., Oswald, W. W., Saenger, C., Anderson, R. S., Bailey, H. L., Clegg, B., Gajewski, K., Hu, F. S., Jones, M. C., Massa, C., Routson, C. C., Werner, A., Wooller, M. J., and Yu, Z. 2016: Holocene climate changes in eastern Beringia (NW North America) – A systematic review of multi-proxy evidence. Quaternary Science Reviews 147, 312–339. https://doi.org/10.1016/j.quascirev.2015.10.021.
Kedrowski, B. L., Crass, B. A., Behm, J. A., Luetke, J. C., Nichols, A. L., Moreck, A. M., and Holmes, C. E. 2009: GC-MS analysis of fatty acids from ancient hearth residues at the Swan Point archaeological site. Archaeometry 51, 110–122.
Lanoë, F. B. and Holmes, C. E. 2016: Animals as raw material in Beringia: Insights from the site of Swan Point CZ4B, Alaska. Am. antiq. 81, 682–696. https://doi.org/10.1017/S0002731600101039.
Lau, H. and Kansa, S. W. 2018: Zooarchaeology in the era of big data: contending with interanalyst variation and best practices for contextualizing data for informed reuse. Journal of Archaeological Science 95, 33–39.
Lepofsky, D. and Armstrong, C. G. 2018: Foraging new ground: documenting ancient resource and environmental management in Canadian archaeology. Canadian Journal of Archaeology/Journal Canadien d’Archéologie 42, 57–73.
Lyman, R. L. 2019: A critical review of four efforts to resurrect MNI in zooarchaeology. J Archaeol Method Theory 26, 52–87. https://doi.org/10.1007/s10816-018-9365-3.
Lyman, R. L. 1984: Bone density and differential survivorship of fossil classes. Journal of Anthropological Archaeology 3, 259–299.
Marshall, F. and Pilgram, T. 1993: NISP vs. MNI in quantification of body-part representation. American Antiquity 58, 261–269.
Martin, A. C. and Barkley, W. D. 1961: Seed identification manual. Univ of California Press.
Martin, P. S. 1984: Prehistoric overkill: The global model. In: P. S. Martin and R. G. Klein (eds.), Quaternary Extinctions. University of Arizona Press, Tucson, AZ, 354–403.
Meier-Augenstein, W. 2002: Stable isotope analysis of fatty acids by gas chromatography–isotope ratio mass spectrometry. Analytica Chimica Acta 465, 63–79.
Meltzer, D. J. 2021: First peoples in a new world: Populating Ice Age America. Cambridge University Press.
Meltzer, D. J. 2000: Renewed investigations at the Folsom Paleoindian type site. Antiquity 74, 35–36.
Mishler, C. 1986: Born with the river: An ethnographic history of Alaska’s Goodpaster and Big Delta Indians (No. DGGS RI 86-14). Alaska Division of Geological & Geophysical Surveys.
Morin, E., Ready, E., Boileau, A., Beauval, C., and Coumont, M.-P. 2016: Problems of identification and quantification in archaeozoological analysis, Part II: Presentation of an alternative counting method. Journal of Archaeological Method and Theory 24, 938–973.
Morisaki, K., Oda, N., Kunikita, D., Sasaki, Y., Kuronuma, Y., Iwase, A., Yamazaki, T., Ichida, N., and Sato, H. 2019: Sedentism, pottery and inland fishing in Late Glacial Japan: a reassessment of the Maedakochi site. Antiquity 93, 1442–1459. https://doi.org/10.15184/aqy.2019.170.
Moss, M. L., Rodrigues, A. T., Speller, C. F., and Yang, D. Y. 2016: The historical ecology of Pacific herring: Tracing Alaska native use of a forage fish. Journal of Archaeological Science: Reports 8, 504–512.
Nelson, R. K. 1983: Make prayers to the raven: a Koyukon view of the northern forest. Chicago: University of Chicago Press.
Neubauer, F. 2018: Use-alteration analysis of fire-cracked rocks. American Antiquity 83, 681–700.
Osgood, C. B. 1937: The ethnography of the Tanaina. Yale University Publications in Anthropology. Yale University, New Haven, CT.
Panshin, A. J. and Zeeuw, C. de 1970: Textbook of wood technology. Volume I. Structure, identification, uses, and properties of the commercial woods of the United States and Canada. Textbook of wood technology. Volume I. Structure, identification, uses, and properties of the commercial woods of the United States and Canada.
Parnell, A., and Govan, E. 2023: simmr: A package for fitting stable isotope mixing models in R. Journal of Statistical Software.
Partlow, M. A. 2006. Sampling fish bones: A consideration of the importance of screen size and disposal context in the North Pacific. Arctic Anthropology 43, 67–79. https://doi.org/10.1353/arc.2011.0064.
Pearson, G. A. 1999: Early occupations and cultural sequence at Moose Creek: A Late Pleistocene site in Central Alaska. Arctic 52, 332–345.
Peres, T. M. 2010: Methodological issues in zooarchaeology. In: A. M. VanDerwarker and T. M. Peres (eds.), Integrating Zooarchaeology and Paleoethnobotany: A Consideration of Issues, Methods, and Cases. Springer, New York, NY, 15–36.
Pitblado, B. L. 2021: On rehumanizing Pleistocene people of the Western Hemisphere. American Antiquity 1–19. https://doi.org/10.1017/aaq.2021.120.
Potter, B. A. 2016: Holocene prehistory of the northwestern subarctic. In: Friesen, T.M., and Mason, O.K (eds.), The Oxford Handbook of the Prehistoric Arctic. Oxford University Press, New York, NY, 537–562.
Potter, B. A. 2007: Models of faunal processing and economy in Early Holocene Interior Alaska. Environmental Archaeology 12, 3–23.
Potter, B. A., Reuther, J. D., Holliday, V. T., Holmes, C. E., Miller, D. S., and Schmuck, N. 2017: Early colonization of Beringia and Northern North America: Chronology, routes, and adaptive strategies. Quaternary International, After Anzick: Reconciling New Genomic Data and Models with the Archaeological Evidence for Peopling of the Americas 444, 36–55. https://doi.org/10.1016/j.quaint.2017.02.034.
Raff, J. A. and Bolnick, D. A. 2014: Genetic roots of the first Americans. Nature 506, 162–163.
Reuther, J. D. 2013: Late Glacial and early Holocene geoarchaeology and terrestrial paleoecology in the lowlands of the middle Tanana Valley, subarctic Alaska. PhD thesis. The University of Arizona, Tucson, AZ.
Richter, K. K., Codlin, M. C., Seabrook, M., and Warinner, C. 2022: A primer for ZooMS applications in archaeology. Proceedings of the National Academy of Sciences 119, e2109323119.
Schoeninger, M. J. 1989: Reconstructing prehistoric human diet. Homo 39, 78–99.
Shaffer, B. S. 1992: Quarter-inch screening: Understanding biases in recovery of vertebrate faunal remains. American Antiquity 57, 129–136.
Shinkwin, A. D. 1979: Dakah De’nin’s village and the Dixthada site. A contribution to northern Athapaskan prehistory. Musée National de l’Homme. Collection Mercure. Commission Archéologique du Canada. Publications d’Archéologie. Dossier Ottawa, 1–197.
Smith, G. M. 2022: The gift of the Middle Tanana: Dene Pre-Colonial history in the Alaskan Interior. Rowman & Littlefield.
Snitker, G., Roos, C. I., Sullivan III, A. P., Maezumi, S. Y., Bird, D. W., Coughlan, M. R., Derr, K. M., Gassaway, L., Klimaszewski-Patterson, A., and Loehman, R. A. 2022: A collaborative agenda for archaeology and fire science. Nature Ecology & Evolution 6, 835–839.
Speth, J. D. 2010: The paleoanthropology and archaeology of big-game hunting: Protein, fat or politics? Interdisciplinary contributions to archaeology. Springer, New York, NY.
Steele, T. E. 2015: The contributions of animal bones from archaeological sites: The past and future of zooarchaeology. Journal of Archaeological Science 56, 168–176.
Stenholm, N. A., Aikens, C. M., and Jenkins, D. L. 1994: Paleoethnobotanical analysis of archaeological samples recovered in the Fort Rock Basin. Archaeological Researches in the Northern Great Basin: Fort Rock Archaeology Since Cressman. Eugene: University of Oregon Anthropological Papers, 531–559.
Stewart, K. M. 1994: Early hominid utilization of fish resources and implications for seasonality and behaviour. Journal of Human Evolution 27, 229–245.
Straus, L. G. 1990: The Early Upper Palaeolithic of Southwest Europe: Cro-Magnon adaptations in the Iberian Peripheries, 40000-20000 BP. In: P. A. Mellars (ed.), The emergence of modern humans: An archaeological perspective. Edinburgh University Press, Edinburgh, Scotland, 276–302.
Surovell, T. A., Allaun, S. A., Crass, B. A., Gingerich, J. A. M., Graf, K. E., Holmes, C. E., Kelly, R. L., Kornfeld, M., Krasinski, K. E., Larson, M. L., Pelton, S. R., and Wygal, B. T. 2022: Late date of human arrival to North America: Continental scale differences in stratigraphic integrity of pre-13,000 BP archaeological sites. PLOS ONE 17, e0264092. https://doi.org/10.1371/journal.pone.0264092.
Surovell, T. A., Pelton, S. R., Anderson-Sprecher, R., and Myers, A. D. 2016: Test of Martin’s overkill hypothesis using radiocarbon dates on extinct megafauna. Proceedings of the National Academy of Sciences 113, 886–891.
Taché, K. and Craig, O. E. 2015: Cooperative harvesting of aquatic resources and the beginning of pottery production in north-eastern North America. Antiquity 89, 177–190.
Thoms, A. V. 2009: Rocks of ages: Propagation of Hot-Rock Cookery in Western North America. Journal of Archaeological Science 36, 573–591.
Thoms, A. V. 2008: The fire stones carry: Ethnographic records and archaeological expectations for Hot-Rock Cookery in Western North America. Journal of Anthropological Archaeology 27, 443–460.
Trigger, B. G. 2006: A history of archaeological thought, 2nd ed. Cambridge University Press, Cambridge, England. Waguespack, N. M. 2012: Early Paleoindians, from colonization to Folsom. The Oxford Handbook of North American Archaeology 86.
Waguespack, N. M. 2005: The organization of male and female labor in foraging societies: Implications for Early Paleoindian Archaeology. American Anthropologist 107, 666–676.
Wygal, B. T., Krasinski, K. E., Holmes, C. E., and Crass, B. A. 2018: Holzman South: A Late Pleistocene archaeological site along Shaw Creek, Tanana Valley, Interior Alaska. PaleoAmerica 4, 90–93.
Yesner, D. R. 1996: Human adaptation at the Pleistocene-Holocene boundary (circa 13,000-8,000 BP) in eastern Beringia. In: L. G. Straus, B. V. Eriksen, J. M. Erlandson, and D. R. Yesner (eds.), Humans at the end of the Ice Age: The archaeology of the Pleistocene-Holocene transition. Plenum Press, New York, NY, 255–276.
Zhilin, M. G. 2014: Early Mesolithic hunting and fishing activities in Central Russia: A Review of the Faunal and Artefactual Evidence from Wetland Sites. Journal of Wetland Archaeology 14, 91–105. https://doi.org/10.1179/1473297114Z.00000000012.