Abstract
Ape knapping experiments complement human knapping experiments as a source of behavioral data to build hypotheses about the learning mechanisms underlying the acquisition of knapping skills in extinct hominins. In addition, ape knapping experiments provide information regarding the stone-related behaviors that could have preceded the systematic production and use of sharp stones in our lineage. In this chapter, we review previous ape knapping experiments with a focus on those that tested apes‘ abilities to socially learn from human demonstrators. Two studies, investigating one orangutan and one bonobo, concluded that both apes could socially learn sharp stone tool-making and use from human demonstrations. These results were interpreted as evidence of the reliance of early hominins on social learning to acquire knapping skills. However, alternative explanations exist. We provide novel data from two experiments investigating the abilities of the two previously untested great ape species (chimpanzees and gorillas) to learn knapping from human demonstrations. Contrary to the previous studies, the chimpanzees and gorillas in our experiments did not acquire sharp stone tool-making or use socially from human demonstrations. However, the apes we tested frequently manipulated the testing materials and two chimpanzees engaged in two events of lithic percussion involving an active hammer (although these actions did not lead to flake detachment). Our results suggest that the observation of human demonstrations is insufficient for the tested apes to acquire knapping abilities. This disparity in results between studies is unlikely to be explained by species differences in tool-use proficiency but rather by the particular rearing background of the previously tested individuals. We discuss how our previous results on both the individual and social learning abilities of unenculturated, untrained orangutans compare to our new results on gorillas and chimpanzees. In addition, we comment on the general implications of ape knapping experiments for understanding the likely origin and maintenance of knapping skills in pre-modern hominins.
Introduction
The production and use of sharp stone tools are often considered one of the most important innovations in human evolution. The use of sharp stone tools allowed our ancestors to access new food sources and engage in a variety of newly available foraging behaviors such as butchering big prey or plant processing techniques (Keeley and Toth 1981; Potts and Shipman 1981). Despite the abundance of sharp stone tools in the archeological record, these artifacts on their own are silent regarding the learning process that naïve hominins underwent to acquire the skills needed for their production and use. Several research avenues, usually involving comparative models, can be pursued in order to investigate this question. One such avenue is to test how modern humans in experimental archaeological studies learn to make and use sharp stone tools (Morgan et al. 2015; Nonaka et al. 2010; Pargeter et al. 2019; Stout et al. 2015; Stout and Semaw 2006). This approach has a long history and presents multiple advantages (Bordes 1969; Eren et al. 2016; Toth 1982, 1985). From a practical perspective, human participants can communicate their perception of their thought processes (e.g., why they are choosing a particular striking platform or hammerstone) as well as receive instructions before and during the experiments (Morgan et al. 2015; Nonaka et al. 2010). From a theoretical point of view, modern humans are the extant species phylogenetically closest to early hominins, potentially allowing to build behavioral and cognitive models of earlier hominin species based on modern human behavior (Stout and Semaw 2006; Toth 1985). However, knapping experiments with humans also have limitations. First, it is often difficult to ensure the naïvety of the human subjects to knapping (as they might have seen movies or museum artifacts) and thus it is difficult to disentangle the origin of knapping skills if they are expressed during the experiment (though see Snyder et al. 2022 for a method used to overcome this limitation using post-test questionnaires and baseline testing). A counterargument to this limitation is that previous knowledge of the task (if any) would most probably involve the final product rather than the knapping technique, meaning that participants would likely be naïve to the production process (though even here, reverse engineering of technique is a possibility and essentially how most lithic analysis must be conducted today given that the production processes are unobservable). The second limitation is that it is often problematic to ensure that other skills in the participant’s repertoire are not being brought to and repurposed for the knapping task (“indirect know-how”; e.g., familiarity with how to break glass, Snyder et al. 2022). However, other behaviors in the early hominin repertoire (e.g., stone throwing, Kühl et al. 2016) could have also preceded and influenced knapping in extinct hominins (Carvalho et al. 2008; Panger et al. 2002), although different learning mechanisms could have underlied their acquisition. The third limitation is that despite being phylogenetically closer to hominins than any other extant primate, the phylogenetic proximity between modern humans and hominins decreases as we explore earlier species. This decrease in relatedness could therefore undermine the use of modern humans as models of earlier hominin species. The fourth limitation of knapping experiments with modern humans is that most modern human populations currently inhabit an environment very dissimilar from the one inhabited by our early ancestors (Faith et al. 2019). In addition, many modern humans have more sedentary lifestyles compared to the nomadic hunter-gathering lifestyle of early hominins (Marlowe 2005). However, such different living conditions between modern and extinct humans could actually be advantageous for experimenters trying to control for previous experience in human knapping studies, as the production and use of stone tools in most modern human populations is rare.
In parallel to human knapping experiments, a research avenue was initiated in the 1970s to investigate the stone tool-making and -using abilities of non-human primates under controlled conditions. Primate knapping studies have the advantage that it is relatively easy to ensure the naïvety of the subjects regarding sharp stone tool-making and -use. Most captive-born primates have been under long-term monitoring and their previous tool-making and -using experiences are known. Primate studies (particularly those involving the genus Pan) can be informative regarding the stone tool repertoires and abilities of older hominin and hominoid species closer to the last common ancestors than our species (Bandini et al. 2022; Carvalho and McGrew 2012; McGrew 2010; Wynn et al. 2011). However, as we will see, primate knapping studies are not free of limitations either.
Prior to our studies, two research projects investigated the abilities of non-human great apes (henceforth apes) to make and use sharp stone tools (several bonobos, Pan paniscus, and one orangutan, Pongo pygmaeus; Toth et al. 1993, 2006; Wright 1972). The goal of these projects was to assess if great apes could acquire sharp stone tool-making and using abilities via the observation of human demonstrations, and if they could, to evaluate the extent of these abilities. The first ape knapping experiment was conducted by Wright (1972) who tested a juvenile male orangutan. The orangutan in Wright’s study (Abang) was provided with a stabilized and pre-shaped flint core fixed on a wooden platform, as well as a hammerstone and an opaque puzzle box baited with food. The lid of the box was kept closed by a rope, which needed to be cut with a sharp object in order to open the lid and thus access the food rewards contained inside the box. This puzzle box represented an indirect task in which the action of cutting with a sharp object did not grant immediate access to the food rewards but instead allowed a door to open through which food could be obtained. In a first experiment, Abang was given demonstrations of how to use a human-made sharp-edged stone as a cutting tool. In addition, Abang experienced at least one instance of molding where the keeper guided Abang’s hand to sever the rope with the stone. These demonstrations and molding were followed by test trials where the orangutan could explore and manipulate the testing materials (i.e., human-made sharp-stones and puzzle box). After nine demonstrations, Abang used a human-made sharp stone as a cutting tool to sever the rope keeping the puzzle box closed. In a second experiment, Wright demonstrated to Abang how to produce sharp stones by using a hand-held hammer to strike on a hand-held core. This knapping technique is known as “free-hand knapping” and it was not actually available to the orangutan during the trials because the core was fixed on a board (see Fig. 1). During the 10th testing day and after 16 demonstrations, Abang produced his own sharp-edged stones by striking a hand-held hammerstone against the fixed core. Abang then proceeded to use one of these sharp-edged stones as a cutting tool to severe the lock-rope, open the baited box and obtain the food rewards (Wright 1972; https://www.youtube.com/watch?v=3exAOxSKYCE).
In the 1990s, Schick, Toth and colleagues similarly tested the language-trained bonobo Kanzi on his abilities to use and produce stone tools after being exposed to demonstrations by a human model (Toth et al. 1993). Additionally, the researchers investigated the development of these skills over a period of several years (Schick et al. 1999; Toth et al. 2006). In the first set of experiments, Kanzi was provided with human demonstrations (before the start of the tests) on how to produce sharp-edged stones using the freehand knapping technique described above as well as on how to use these sharp-edged stones as cutting tools to sever a rope (Toth et al. 1993). After having been exposed to these demonstrations, Kanzi was given hammerstones, loose stones of different sizes and raw materials (although eventually, only chert cobbles were used in the tests), and a puzzle box with a rope-lock similar to the one used by Wright baited with food (see above). In later experiments, Kanzi was also provided with a second “drum-like” puzzle box. This second box consisted of a wooden cylinder covered by a transparent plastic lid that needed to be cut in order to (directly) access the food placed inside the cylinder. On the first day of the first stone tool-use experiment, after having observed human demonstrations of stone tool-production and -use, Kanzi started to use human-made flakes as cutting tools to sever the rope-lock of the baited puzzle box. In a follow-up experiment, Kanzi’s abilities to produce sharp-edged stone tools after observing human demonstrations of the freehand knapping technique were evaluated. After a month of trials and several unsuccessful attempts at producing sharp-edged stones using the freehand knapping technique, Kanzi developed his own technique to initiate stone fracture where he threw a cobble against a hard surface (“throwing technique,” Fig. 1). This innovative solution was not the type of solution originally intended by the experimenters and so attempts were made to discourage this behavior by moving the experiments to an outdoor enclosure with soft ground. Additionally, he also innovated a variant of the throwing technique where he threw a cobble against another stationary cobble (“directed throwing technique,” Fig. 1). However, the timing and context of this innovation is not described (Toth et al. 1993). Eventually, Kanzi also successfully performed the demonstrated freehand technique. In later years, Kanzi preferably used the two throwing techniques over the freehand knapping technique demonstrated to him by humans (Schick et al. 1999). Kanzi’s half-sister Panbanisha was later reported to have learnt to use and produce sharp-edged stone tools via freehand percussion after observing a human demonstrator (Savage-Rumbaugh and Fields 2006). Similarly, Panbanisha’s two sons were also reported to have acquired sharp-edged stone-making and -using skills via the observation of the two more experienced bonobos (Kanzi and Panbanisha), although neither the learning process nor the bonobos’ knapping skills were tested nor described in detail (Toth et al. 2006) and should therefore be considered with caution.
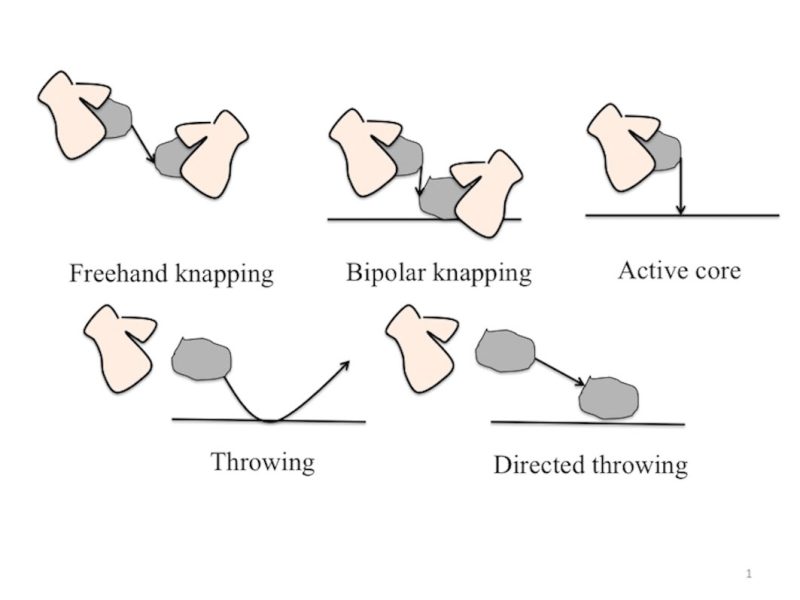
Despite being highly innovative at the time, the ape knapping studies described above present a series of methodological limitations in light of recent developments in animal cognition research. Specifically, all individuals tested in these previous studies were highly- or at least semi-enculturated, meaning that they were partly raised in a human cultural environment that included extensive direct human contact and intentional training (Furlong et al. 2008; Henrich and Tennie 2017). This is problematic for making inferences based on the results of these earlier studies. Enculturated apes cannot be considered representative models of their wild counterparts, nor of their last common ancestors with the hominin lineage (Henrich and Tennie 2017). Indeed, it is widely accepted that one of the tested apes (the bonobo Kanzi) represents one of the most extreme cases of such human enculturation (Savage-Rumbaugh et al. 1986). As for Abang the orangutan, a former ape keeper from the zoo where the experiments took place told us (AMR) in 2019 that it was common practice in the 1960s and 1970s for the keepers in Bristol Zoo (UK, where Abang was located at the time) to enter the enclosure with the orangutans as well as to take them for walks around the zoo grounds. Such close contacts and frequent human interactions (forbidden in most zoos today) suggest that Abang the orangutan was at least semi-enculturated (Henrich and Tennie 2017). In Abang’s case, there is the additional limitation that molding took place during the first experiment exploring sharp stone tool-use. Abang’s keeper physically guided Abang’s hands to sever the rope-lock using a sharp stone. This instance of molding confounds the results of Abang’s learning process of sharp stone tool-use (though note that Wright did not report any molding taking place during or before the sharp-stone tool-making experiments). Relatedly, cobbles were actively placed into Kanzi’s hands to promote stone tool-making (Savage-Rumbaugh and Fields 2006). Finally, the reduced sample sizes (N=1) in both studies (Toth et al. 1993; Wright 1972) leave open the question of the generalizability of their results to other ape subjects of the same and other species (though some limited data exist for three more bonobos, see above).
In the present chapter, we expand on the findings of these early experiments, first, by testing the two remaining species of great apes, namely chimpanzees and gorillas, and, second, by addressing some of the limitations and confounds in these early ape experiments. We investigated—for the first time—whether group-housed chimpanzees and gorillas can acquire sharp stone tool-making and -using abilities via the observation of human demonstrations. We tested a group of chimpanzees including individuals both with limited levels of enculturation (semi-enculturated) and unenculturated as well as a group of unenculturated gorillas. Here, we used the term semi-enculturated as defined by Henrich and Tennie (2017) to refer to apes that were hand-reared while they were young but then lived most of their lives in a conspecific group at a zoological institution. By unenculturated individuals we refer to captive individuals which have always lived in conspecific groups, are mother-reared and have not received extensive human training (e.g., except for veterinary procedures). We ensured the naïvety of the test subjects to the target tasks by interviewing the ape keepers and confirming the lack of previous stone tool-making and -using experience of the apes both within and outside previous experiments. If naïve, unenculturated, untrained chimpanzees and gorillas would develop sharp stone tool-making and -using abilities following human demonstrations (but not on their own), this would mean that the results of Wright and Toth et al. might be generalizable to all great ape species. In turn, such findings would suggest that human demonstrations suffice for the acquisition of these abilities even in naïve, untrained, unenculturated individuals. Such results would also provide support for the hypothesis that the earliest stone tool-using hominins learnt stone tool-making and -using abilities via the observation of other individuals’ behavior. If naïve, unculturated and untrained chimpanzees and gorillas would not express the target behaviors after observing human demonstrations, this would suggest that human training and/or enculturation, perhaps in addition to human demonstrations, are required for the acquisition of these behaviors in these species (compare Bandini, Motes-Rodrigo et al. 2021, 2022). Such findings would suggest that the background and training of the bonobos and orangutans tested in the previous ape experiments had a strong influence on the positive findings observed (causing enhanced innovativeness and/or increased social learning abilities).
Materials & methods
Subjects and housing
The subjects of the experiments presented here were gorillas (Nadults = 2F and 2M; Njuvenile = 1M; Ninfant = 1M; mean age = 23.5 years ± 16) and chimpanzees (Nadults = 7F and 6M; mean age = 33 years ± 11) housed at Twycross Zoo (Atherstone, UK). The chimpanzees were housed in a group that included seven human-reared, three mother-reared and three individuals of unknown rearing. The gorillas were housed in a group that included four mother-reared individuals and two individuals of unknown rearing. All these apes had access to indoor and outdoor enclosures as well as to quarters off-sight from the visitors. All ape indoor enclosures were equipped with environmental enrichment such as climbing frames, bedding materials, platforms and containers where food could be placed for the apes to retrieve. The floor of the indoor enclosures was covered with wooden chips and straw. The apes’ outdoor enclosures consisted of grassed areas surrounded by glass walls from where visitors could observe the apes. The outdoor enclosures included climbing frames and huts. Feedings took place several times a day when food (fruit, vegetables, primate pellets and nuts) was scattered in the indoor and outdoor enclosures. Food was often placed inside enrichment devices such as hanging balls and boxes attached to the meshes. Water was available ad libitum in all enclosures. The experiments took place in the off-sight quarters connected to the indoor enclosures. During the experiments, all apes could access the off-sight quarters as well as both the indoor and outdoor enclosures as a group.
Testing materials
Both during the demonstrations and the tests we used two puzzle boxes (the tendon box and the hide box), three artificial hammerstones fixed to the enclosure bars using chains and a fixed chert core. The tendon box was modeled on an earlier version described by Wright (1972) and Toth et al. (1993) and consisted of two opaque boxes secured to a wooden board (Fig. 2). The tendon box had a clear Plexiglas window (5 cm x 16 cm) at the top that allowed the apes to see the reward inside one of the boxes. The door of the reward box was pulled shut by a rope that ran through the inside and exited through a hole in the opposite end. The rope then ran between the two boxes for approximately 5cm and entered the second (non-rewarded) box. Thus, the rope was only accessible in the area between the two boxes and had to be cut there to allow the door of the reward box to open.

The hide box was designed based on an apparatus used to test capuchin monkeys (Cebus apella) (Westergaard and Suomi 1994) and consisted of a transparent Plexiglass cylinder (16 cm wide x 15.5 cm high) with a metallic rim (Fig. 3). A silicone membrane 2 mm thick was screwed in between the cylinder and the rim, blocking the access to the reward placed inside the cylinder. Three artificial hammers were made out of concrete and used during demonstration tests (small: 12 cm length x 9 cm width, 2 kg; medium: 15 cm length x 10 cm width, 2.5 kg; large: 18 cm length x 11 cm width, 3 kg). The hammers had an overall potato shape and were built around a metallic scaffold linked to a chain that allowed fixing the hammers to a wooden platform. The hammers were tested a priori on cores equivalent to the ones used in the experiments to ensure that they allowed flake removal from the cores. The concrete used to build the hammers included particles of up to 0.5 cm in diameter (Fig. 3). Retouched Norfolk Chert cores were used for the demonstrations and tests. The cores were modified to display angle variability between ~90 degrees and ~40 degrees. Different cores were used for the demonstrations and the tests. If a core was not modified during a test, the core was used in further tests. Due to safety regulations, the core had to be fixed on a metallic platform (20 cm x 20 cm x 2 cm) to prevent the apes from carrying the core into the indoor enclosure (similar to Wright 1972). The core was attached to the platform using a metallic wired mesh with a hole width of 50 mm and wire diameter of 3 mm from XTEND (Carl Stahl ARC GmbH, Architectural Cables and Mesh Systems). This fixing system left exposed a portion of the core (Fig. 3). The core was attached to the platform ensuring that the acute angle from which flakes could be detached was facing up.


Experimental procedure
Demonstrations to all apes were made in a group setting. The demonstrations to the gorillas took place through a glass wall in the indoor enclosure while the sleeping quarters were being cleaned and before the zoo opened to the public. The demonstrations to the chimpanzees took place through a mesh in the service aisle in front of the off-sight quarters where the tests took place and which the chimpanzees were free to access outside of cleaning hours (see Fig. 10 in Neadle et al. 2020). An individual was considered to have observed a demonstration when his/her head was oriented towards the demonstrator (AMR) during the entire demonstration. If the individual moved away or stopped looking during the demonstration, the demonstrator stopped and started again from the beginning once the individual was again paying attention. A spreadsheet of which individuals had observed which demonstrations was continuously updated after each demonstration (this was necessary to ensure that the chimpanzees saw a minimum of three demonstrations before their first test and to count how many demonstrations each gorilla saw, see below). The identity of the individuals that observed each demonstration was confirmed by the keepers present during the demonstrations. If the experimenter was not sure if an individual had seen a full demonstration, it was assumed that he/she had not and the demonstration was repeated. During all demonstrations, the wooden platforms where the testing materials were fixed were placed on the floor between the apes and the demonstrator, so the actions of the demonstrator were clearly visible from where the apes were.
Each demonstration consisted of the production of one flake by striking the stabilized core on the fixing platform with one of the artificial hammers. This was followed by the use of the produced flake to open one of the puzzle boxes and obtain the food reward. This knapping technique was employed in order to demonstrate to the apes the flake production method that later was going to be available to them during the tests (unlike Wright 1972). Only one flake was made in each demonstration and flakes were not reused between demonstrations. After detaching a flake, the demonstrator held it in front of the observing apes to ensure that he/she had seen the flake. The subsequent demonstrations of flake use did not start until all apes present had seen the flake (i.e., their head was oriented towards the demonstrator while she was holding the flake). Demonstrations of flake use were conducted with both puzzle boxes (see below). When demonstrating how to open the tendon box, the demonstrator used the flake she had produced immediately before to cut the rope that kept the door of the box closed. When demonstrating how to open the hide box, the demonstrator used a flake she had produced immediately before to cut through a plastic sheet placed in the same position as the silicone membrane would be placed during the actual tests. We used plastic sheets instead of silicone membranes during the demonstrations due to the limited availability of silicone membranes and the high number of demonstrations. When obtaining the reward, the demonstrator made sure that the ape saw it by taking the reward out of the box and showing it to the observing apes. After each demonstration, the boxes were re-baited with the same reward and closed. Cores used during the demonstrations were exchanged for new cores before the apes had access to the testing materials.
The demonstrations involved all possible combinations (N=9) between hand (left, right, both) and hammer type (small, middle, large). Each of the nine combinations was demonstrated twice per test box (3 hand combinations x 3 hammers x 2 boxes x 2 rounds of demonstrations = 36 demonstrations) before the start of Test 1. As all demonstrations had to be made in the presence of a keeper to comply with zoo regulations, each round of demonstrations was spread over at least two days, depending on the keeper’s availability. A maximum of two cores were used per demonstration day and these were exchanged when their knappable surfaces were exhausted.
For the gorillas, the experiments were structured into the following phases:
Initial Demonstrations (N=36) – Test 1 – Test 2 – Repeated Demonstrations (N=12) – Test 3 – Test 4. Each gorilla saw at least six demonstrations before the first test.
For the chimpanzees, the experiments were structured into the following phases:
Initial Demonstrations (N=36) – Test 1 – Test 2 – Repeated Demonstrations (N=18) – Test 3 – Test 4 – Test 5 – Test 6. As some of the chimpanzees rarely entered the off-sight quarters where demonstrations were taking place, we proceeded to Test 1 when at least 80% of the chimpanzees had seen a minimum of three demonstrations (Table 1). Tests 5 and 6 were implemented given the results of Tests 1 and 3.
Repeated demonstrations were implemented after Test 2 to account for any potential effects of the delay since the initial demonstrations. These demonstrations were meant to act as reminders of the task and solutions. Each test took place on a consecutive day and the testing materials were only available to the apes during the tests. Apes participated in a maximum of one test per day, which lasted for several hours (between 2 and 3 hours in the case of chimpanzees and approximately 2h in the case of gorillas) on a voluntary basis. The testing materials (including the baited boxes) were placed inside the testing quarter before each test as described above (Fig. 3) and all individuals were free to participate. All demonstrations and tests were recorded with two Sony HDR-CX330E Handycams video cameras. From the video recordings of each test, the demonstrator later coded all active interactions that the chimpanzees and the gorillas performed with the testing materials. An interaction started when the ape entered in physical contact with the testing materials and finished when the contact a) ceased, b) paused for more than three seconds or c) the interaction type changed (the part of the materials explored changed). We only considered active interactions, meaning that stationary contact (such as placing and resting a hand on the materials, sitting or lying down on the materials) was not coded. From each interaction we coded: 1) the identity of the individual; 2) the testing material that the individual interacted with (core, flake, hammer, hide box, tendon box); 3) if the interaction took place manually, using the mouth or a tool; 4) the type of tool and 5) the duration of the interaction.

Results
Chimpanzees
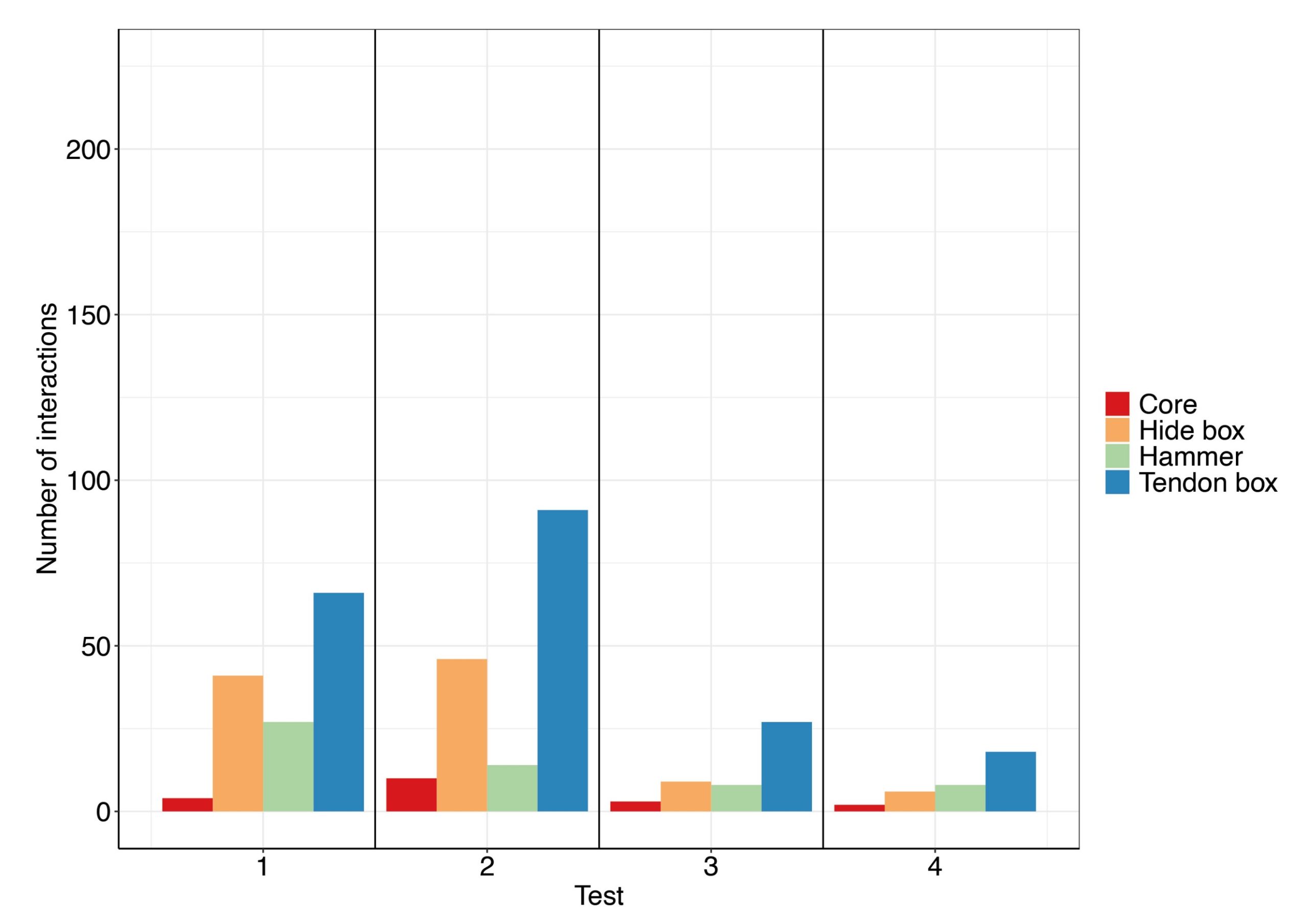
Twelve out of the 13 chimpanzees tested interacted at least once with the materials during the tests. The total number of interactions was 1025. The total number of interactions per individual varied from two to 199 (mean number of interactions ± SD= 85 ± 65) and the number of interactions per test varied from 473 during Test 1 to 68 during Test 5 (Fig. 4). The chimpanzees interacted the most with the baited boxes (hide and tendon box, Fig. 5). One of the chimpanzees learnt to open the hide box manually by pulling on the edge of the silicone membrane and did so in several tests. The manual opening of the box sometimes took place when other individuals were present in the testing quarter. No other chimpanzee was successful in opening any of the baited puzzle boxes. Despite being empty after it had been manually opened, most chimpanzees interacted with the hide box (including the individual that learnt how to open it manually, Fig. 5), suggesting that the food rewards were not the only motivator of the chimpanzees’ exploratory behavior.

Most of the interactions performed by the chimpanzees with the testing materials were manual (N=970), although the chimpanzees also used their mouths to try to open the boxes (N=5) as well as several tools that they brought from the indoor enclosure (N=50). The chimpanzees performed 20 interactions using sticks and five using pieces of straw obtained from the indoor enclosure. The nature of the tools used in the remaining 25 interactions could not be identified from video recordings.
Twelve chimpanzees touched the rope of the tendon box a total of 52 times. Of these, only two individuals (CH3 and CH11) interacted with the rope more than five times (13 and ten times, respectively). Most interactions with the rope took place by hand (N=45) and using the teeth (N=5), but on two occasions a piece of straw was used to apparently try to (unsuccessfully) break the rope. The straw pieces were looped around the rope of the tendon box and used to pull the rope upwards.

The chimpanzees interacted with the hammers a total of 155 times. Of these interactions, 12 involved percussion, defined as the use of tools to strike surfaces or objects (Whiten et al. 2009). On eight of these occasions, the chimpanzees used a hammer to hit the wooden platform holding the core and hammers (CH2: M, 33 years old at the time of testing, human-reared; CH7: F, 31, human-reared; CH12: M, 40, rearing unknown). On four of these occasions, one chimpanzee used a hammer to hit another hammer (CH12). Two chimpanzees also forcefully rolled the hammers on their side without lifting them (N=6; CH2 and CH7), which caused a hammer to impact with the core. Eight times, two chimpanzees (CH7 and CH12) made some contact with the core with a (hand-held) hammer. Two of these interactions involved percussion. CH7 was the first individual to use the small hammer to hit the middle section of the core once during Test 1. During Test 3, CH12 struck four times the core with the middle-sized hammer targeting the mesh-covered section of the core (Fig. 6). None of these interactions led to flake detachment as the strength employed to strike the core was insufficient and the target of the percussive actions was not the exposed acute angle of the core.

Gorillas
No attempt at stone tool-making was ever observed in the gorillas. All six gorillas interacted at least once with the materials provided. The total number of interactions was 380. The total number of interactions per individual ranged from eleven to 208 and the number of interactions per test varied from 34 on Test 4 to 161 on Test 2 (Fig. 7). The gorillas interacted the most with the baited boxes (hide and tendon box, Fig. 8) followed by the hammers and lastly the core.

Most of the interactions performed by the gorillas with the testing materials were manual (N=341), although the gorillas also used their mouths to seemingly try to open the boxes (N=19). In addition, the gorillas also used several sticks as probing tools to interact with the boxes that they brought from the indoor enclosure (N=20). Four of the gorillas touched the rope of the tendon box a total of 41 times (across subjects). Of these, only two individuals (the juvenile and the infant) interacted with the rope more than five times (22 and 17 times, respectively). Most interactions with the rope took place by hand (N=26) and using their teeth (N=14), and on one occasion a stick was pressed down against the rope.
The gorillas did not engage in any type of percussive activity that involved the hammers. However, they did play with the hammers by rolling them on the platforms eleven times in total across individuals. In Tests 1, 2 and 4, one adult female managed to open the hide box by hand by pulling on the silicone and ripping it. The manual opening of the box sometimes took place when other individuals were present in the testing quarter. Other individuals nevertheless explored the empty box frequently even though it was empty. No other gorillas were successful in opening any of the two puzzle boxes.
Discussion
Two out of 13 tested chimpanzees (both of whom were potentially enculturated) but no gorilla out of six tested used a hand-held artificial hammer to strike a fixed core in a variant of the bipolar technique. These striking actions performed by the chimpanzees took place after observing human demonstrations of how to make and use sharp stone tools. These two instances of percussion were extremely rare among the total of 1025 interactions observed in this chimpanzee group. No sharp-edged stone detachment took place as a consequence of these two striking actions. This lack of stone detachment was probably caused by the use of insufficient force by the chimpanzees. Additionally, in these two instances of percussion, the chimpanzees did not target the exposed area of the core that had suitable angles for knapping, but instead hit the center of the core (which was covered by a metal mesh).
The lack of tool use in wild populations alone would not explain the negative results of our experiments with captive gorillas. Although gorillas generally do not use tools in the wild (though see Breuer et al. 2005), they have been reported to use tools with similar proficiency as other apes in captivity (Shumaker et al. 2011). On the other hand, chimpanzees are the great ape species with the most varied tool-use repertoire in the wild and the only species of ape (so far) that uses stones as tools in the wild (Biro et al. 2006; Boesch and Boesch 1984). Stone tool use (including percussion) in foraging contexts has been described in multiple populations of wild chimpanzees (Carvalho et al. 2008; Koops et al. 2010; Luncz et al. 2012) but this behavior is relatively rare or absent among captive, untrained individuals even when stones are provided as testing materials (Arroyo et al. 2016). A previous study investigating the spontaneous abilities of chimpanzees to acquire sharp stone tool-making and using skills during baseline tests (in the absence of demonstrations) showed that captive, unenculturated chimpanzees from two different groups do not spontaneously engage in lithic percussion (Bandini, Motes-Rodrigo, et al. 2021).
Although rare, the two observations of lithic percussion collected during the present study could indicate that the chimpanzees acquired some action information regarding bipolar knapping from the demonstrations (i.e., know-how copying). However, alternative explanations are also possible. Given the published literature on chimpanzees’ lack of spontaneous know-how copying (Motes-Rodrigo et al. 2021; Neadle et al. 2020; Tennie et al. 2012), the two chimpanzees in our study that engaged in stone percussion could have socially learnt from the demonstrations not the know-how of the underlying behavior, but the target of percussion (know-where/what). The social learning mechanism underlying this acquisition of information would thus have been stimulus and/or local enhancement (acquisition of know-what and/or know-where). The use of the artificial hammers as active elements could be then explained by the fact that these were the only mobile materials available in the testing quarter (i.e., the know-what to use as hammers was a given).
Of the two chimpanzees that performed percussive actions targeted towards the core, one of them (CH7) had been hand-reared by keepers as an infant and the other (CH12) had an “unknown background.” It is therefore possible that both (but at least one) of the two chimpanzees that performed percussive activities were somehow enculturated due to extensive exposure to human contact during an extended period of time (Furlong et al. 2008; Henrich and Tennie 2017). This is problematic for making phylogenetic and species-wide inferences. Extensive human contact has been shown to affect both brain connectivity and cognition—including motivation and/or ability to copy behavioral forms or action know-how (Pope et al. 2018; Tomasello et al. 1993; Tomasello and Call 2004). Enculturated apes have been repeatedly shown to possess enhanced copying abilities compared to unenculturated apes (including know-how copying, Call 2001; Call and Tomasello 1996; Custance et al. 1995). Indeed, the only two studies that claimed that great apes could acquire sharp-edged stone tool-making skills (by copying a human model) tested enculturated or semi-enculturated individuals who had extensive human training and contact throughout their lives (Toth et al. 1993; Wright 1972). Therefore, enhanced attention towards humans and/or the enhancement/training of certain cognitive abilities (such as know-how copying, compare Heyes 2018) via enculturation could explain the positive results of earlier ape knapping studies (see introduction) and the two observations of chimpanzee lithic percussion in the present study. However, if this were the case, it remains unanswered why the other hand-reared individuals in the group did not perform percussive activities after being exposed to demonstrations. A potential explanation for this latter observation could be that a combination of human exposure during rearing, a more neophilic personality or differences in motivation levels (Forss et al. 2020; Hopper et al. 2014) led only two individuals to perform percussive actions (generally, animal behavior remains to some degree stochastic, even after training and enculturation). Testing hand-reared (and especially enculturated) individuals also incurs some of the limitations of human knapping studies highlighted in the introduction section. For instance, it might be difficult to exclude that hand-reared and/or enculturated apes are naïve to behaviors or materials of interest. In such cases, enculturated (but not unenculturated apes, Clay and Tennie 2017) might apply a previously known behavior to a new context, material or task after seeing a demonstration (i.e., contextual imitation). However, the hand-reared individuals in our study were hand-reared at zoological institutions and after interviewing the zoo keepers we could confirm their naïvety towards the testing materials and target behaviors.
Yet, even if we were to consider both chimpanzees that engaged in lithic percussion in this study as enculturated, our subjects and the subjects tested in previous ape knapping studies (possibly Abang, but certainly Kanzi) would clearly differ in their degree of enculturation. Kanzi the bonobo is one of the most famous cases of an ape brought up in a human social environment and exposed to language experiments throughout his life. Kanzi is renowned for having received extensive training to communicate with humans using lexigrams (Savage-Rumbaugh et al. 1986) and was accustomed to being in the same room as researchers and interacting with them. Although the orangutan tested by Wright (Abang) was not specifically nor intentionally trained to communicate with humans or copy them, he was often in very close contact with humans as reported by a keeper of the ape team at the time of the experiments conducted by Wright (which can provide unintentional training across domains). Some of the chimpanzees we tested here were reported to be hand-reared and other subjects (including gorillas) had unknown backgrounds. However, it is unclear which degree of enculturation those apes would have as it is not described what hand-rearing involved or during how long (if at all) the hand-reared chimpanzees (and perhaps some gorillas) were separated from their group and in close, direct contact with humans. In any case, the chimpanzees included in our study would have only lived in close proximity to humans during their infancy as they were introduced into the conspecific group when they were juveniles. Consequently, their degree of enculturation would be lower (semi-enculturated) than that of Abang or Kanzi (enculturated), who lived in human-cultural environments even in adulthood. This difference in enculturation degree could partly explain the different results obtained between studies, suggesting that an elevated level of enculturation (perhaps including extensive human exposure) is necessary for apes to acquire sharp stone tool-making and use – either on their own or triggered/copied from human demonstrations (compare Motes-Rodrigo et al. 2022).
The results presented here also contrast with one of our previous studies testing the social learning abilities of three stone-tool naïve, untrained and unenculturated orangutans to learn from human demonstrations how to make and use sharp-stone tools (Motes-Rodrigo et al. 2022). In this previous experiment, we employed a similar testing set-up and equivalent materials as those employed in the present study. In the orangutan experiment, one young female (O1) engaged in frequent percussive behavior (N=19 events) using a hand-held hammer to strike a platform holding the hammers, the walls of the enclosure and the fixed core. In these core-directed strikes, and contrary to the chimpanzees we tested here, the target of percussion was the exposed area of the core (i.e., the only region from which sharp stones could have been detached). In addition, percussive actions of the orangutan using the hammers to strike against the walls and holding platforms led to the detachment of several pieces from the hammers, some of which were sharp enough to be used as cutting tools. Nevertheless, the orangutan did not use these pieces as tools. Previous experiments included in that same study on a different orangutan population showed that orangutans spontaneously engage in lithic percussion during baselines without the need to observe human demonstrations of percussion. These spontaneous percussive events involved the use of both hammers and the core as active elements to strike the walls and floor of the testing quarter. In one trial, the core was used as an active element when an individual removed it from its fixing platform. This passive hammer technique1 led to the detachment of three sharp stone fragments. No observations were made of orangutans spontaneously using the hammers to strike the core in the absence of human demonstrations. Given this spontaneous, untrained expression of lithic percussion by naïve orangutan individuals (see also examples in other orangutan populations Bandini, Grossmann, et al. 2021; and capuchins Westergaard and Suomi 1994) it is likely that O1 did not acquire the know-how involved in lithic percussion from the demonstrations but instead socially gathered and used information regarding the target (know-where/know-what) of percussion.
The results of the present and previous studies on ape knapping point to several factors influencing the acquisition process of sharp stone-tool production by apes. Here, stone tool-making or production should be understood as the general ability to detach sharp pieces from a core or from a (human-intended) hammerstone rather than the production of sharp artifacts by specifically striking a platform-oriented at an acute angle relative to a high mass zone (acute angle rule, Moore 2020), for which there is no evidence in apes. A combination of enculturation, training/molding and the provision of human demonstrations might allow some apes (at least one bonobo and one orangutan) to reliably acquire sharp stone tool-making and -using abilities during individual tests over extended periods of time. This combination of subject characteristics and testing methodology seems to be required for apes to learn these skills considering the results presented here. When hand-reared chimpanzees (in conspecific groups with unenculturated individuals) were provided with human demonstrations, the observed stone-related behaviors were much rarer and limited to only certain actions (i.e., occasional percussion) that are not outside the spontaneous abilities of naїve individuals (Bandini, Grossmann, et al. 2021; Motes-Rodrigo et al. 2022). In orangutans, it seems that human demonstrations (provided in an individual setting) are not sufficient for the acquisition of the complete sequence of stone tool-making and -use when the individuals are not enculturated (Motes-Rodrigo et al. 2022). Thus, one potential explanation for the different results obtained in the present compared to previous studies with positive results (Toth et al. 1993; Wright 1972) is the specific degree of enculturation. Unenculturated apes can sometimes show some elements of the knapping sequence and even sharp stone tool use (Motes-Rodrigo et al. 2022) but these skills are shown rarely and unreliably, and so far by single individuals.
Overall, ape knapping experiments suggest that elevated levels of enculturation—perhaps in addition to human demonstrations2 and/or training— are required for apes to reliably express stone tool-making skills. In addition, modern task-naïve, unenculturated and untrained apes cannot individually or socially learn the complete sequence of behaviors involved in knapping, including the use of the resulting sharp stones. However, some of these experiments have revealed that at least some species of apes can spontaneously acquire some elements of this sequence (lithic percussion and the use of a sharp stone as a cutting tool, separately, Motes-Rodrigo et al. 2022). These results open the possibility that our hominid ancestors possessed the anatomical and cognitive abilities to engage in and learn certain behavioral prerequisites of lithic technologies in the absence of know-how copying. For example, hominids could have unintentionally produced sharp-edged stones as a by-product of percussive activities (e.g., during foraging, play or another activity, as it occurs in other species; Proffitt et al. 2016). In addition, it is possible that they could also have spontaneously used flakes available from anthropogenic (unintentional sharp-edged stones produced as a by-product of percussive activities; perhaps even by other species) and/or flakes and/or unmodified stones from non-anthropogenic origins (e.g., cliff falls, Barnes 1939) as cutting tools (e.g., Mountford 1941). Later hominins could have then combined these abilities leading to the emergence of intentional knapping.
Despite these and other advances in non-human and human primate archaeology, the emergence process of the complete hominin knapping sequence, including intentional sharp-edged stone production and use, remains speculative. Similarly, it is unclear whether intentional knapping evolved as a response to necessity, opportunity, relative profitability or a combination of these (Koops et al. 2014; Rutz and St Clair 2012). Knapping skills could have emerged in our lineage when the degree of terrestriality increased and the foraging niche expanded (Koops et al. 2014), including the exploitation of larger prey and processing of hard underground storage organs of plants (Marchant and McGrew 2005), which could have increased the need or relative profitability of sharp stone tools (Shea 2011). We are hopeful that future investigations will better be able to determine both why and when these skills emerged in our ancestors. However, even if we find the answers to these questions, the puzzle will remain of when and why the acquisition of sharp stone-tool production and use started to depend on know-how copying skills (Bandini, Motes-Rodrigo, et al. 2021; Motes-Rodrigo et al. 2022; Snyder et al. 2022; Tennie et al. 2016, 2017).
2 Please note that baseline tests (in the absence of demonstrations) with highly unculturated apes are still pending.
Acknowledgments
This project received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (grant agreement number 714658; STONECULT project). The authors are also grateful to Dr. Ammie Kalan and one anonymous reviewer for their valuable comments on previous versions of this chapter.
References
Arroyo, A., S. Hirata, T. Matsuzawa, and I. de la Torre. 2016. Nut Cracking Tools Used by Captive Chimpanzees (Pan troglodytes) and Their Comparison with Early Stone Age Percussive Artefacts from Olduvai Gorge. PLoS One 11: e0166788.
Bandini, E., J. Grossmann, M. Funk, A. Albiach-Serrano, and C. Tennie. 2021. Naïve orangutans (Pongo abelii and Pongo pygmaeus) individually acquire nut-cracking using hammer tools. American Journal of Primatology 83(9): e23304. DOI: 10.1002/ajp.23304.
Bandini, E., R.A. Harrison, and A. Motes-Rodrigo. 2022. Examining the suitability of extant primates as models of hominin stone tool culture. Humanities and Social Sciences Communications 9(1): 74. DOI: 10.1057/s41599-022-01091-x.
Bandini, E., A. Motes-Rodrigo, W. Archer, T. Minchin, H. Axelsen, R.A. Hernandez-Aguilar, S.P. McPherron, and C. Tennie. 2021. Naïve, unenculturated chimpanzees fail to make and use flaked stone tools [version 1; peer review: 3 approved]. Open Research Europe 1: 20. DOI: 10.12688/openreseurope.13186.1.
Barnes, A. S. 1939. The differences between natural and human flaking on prehistoric flint implements. American Anthropologist 41: 99–112.
Biro, D., C. Sousa, and T. Matsuzawa. 2006. Ontogeny and cultural propagation of tool use by wild chimpanzees at Bossou, Guinea: Case studies in nut cracking and leaf folding. In Cognitive development in chimpanzees, ed. by T. Matsusaka, M. Tomonaga, and M. Tanaka. Springer.
Boesch, C., and H. Boesch. 1984. Possible causes of sex differences in the use of natural hammers by wild chimpanzees. Journal of Human Evolution 13(5): 415–440. DOI: 10.1016/s0047-2484(84)80055-x.
Bordes, F. 1969. The Corbiac blade technique and other experiments. Tebiwa 12: 1–22.
Breuer, T., M. Ndoundou-Hockemba, and V. Fishlock. 2005. First Observation of Tool Use in Wild Gorillas. PLoS Biology 3(11): e380. DOI: 10.1371/journal.pbio.0030380.
Call, J. 2001. Body Imitation in an Enculturated Orangutan (Pongo Pygmaeus). Cybernetics and Systems 32(1–2): 97–119. DOI: 10.1080/019697201300001821.
Call, J., and M. Tomasello. 1996. The effect of humans on the cognitive development of apes. In Reaching into thought: The minds of the great apes, ed. by A. E. Russon, K. A. Bard, and S. T. Parker, pp. 371–403. Cambridge University Press.
Carvalho, S., E. Cunha, C. Sousa, and T. Matsuzawa. 2008. Chaînes opératoires and resource-exploitation strategies in chimpanzee (Pan troglodytes) nut cracking. Journal of Human Evolution 55: 148–163. DOI: 10.1016/j.jhevol.2008.02.005.
Carvalho, S., and W.C. McGrew. 2012. The origins of the Oldowan: Why chimpanzees (Pan troglodytes) still are good models for technological evolution in Africa. In Stone Tools and Fossil Bones: Debates in the Archaeology of Human Origins, ed. by M. Domínguez-Rodrigo, pp. 222–244. Cambridge University Press.
Clay, Z., and C. Tennie. 2017. Is Overimitation a Uniquely Human Phenomenon? Insights From Human Children as Compared to Bonobos. Child Development 89: 1535–1544. DOI: 10.1111/cdev.12857.
Custance, D. M., A. Whiten, and K.A. Bard. 1995. Can Young Chimpanzees (Pan troglodytes) Imitate Arbitrary Actions? Hayes & Hayes (1952) Revisited. Behavior 132: 837–859.
Eren, M. I., S. J. Lycett, R.J. Patten, B. Buchanan, J. Pargeter, and M.J. O’Brien. 2016. Test, Model, and Method Validation: The Role of Experimental Stone Artifact Replication in Hypothesis-driven Archaeology. Ethnoarchaeology 8(2): 103–136. DOI 10.1080/19442890.2016.1213972.
Faith, J. T., J. Rowan, and A. Du. 2019. Early hominins evolved within non-analog eco-systems. Proceedings of the National Academy of Sciences 116(43): 21478–21483. DOI: 10.1073/pnas.1909284116.
Forss, S. I. F., A. Motes-Rodrigo, C. Hrubesch, and C. Tennie. 2020. Chimpanzees’ (Pan troglodytes) problem-solving skills are influenced by housing facility and captive care duration. PeerJ 8: e10263.
Furlong, E. E., K.J. Boose, and S.T. Boysen. 2008. Raking it in: The impact of enculturation on chimpanzee tool use. Anim Cogn 11: 83–97. DOI: 10.1007/s10071-007-0091-6.
Henrich, J., and C. Tennie. 2017. Cultural Evolution in Chimpanzees and Humans. In Chimpanzees and Human Evolution, ed. by M. Muller, R. Wrangham, and D. Pilbeam, pp. 645–702. Harvard University Press.
Heyes, C. 2018. Cognitive gadgets: The cultural evolution of thinking. Harvard University Press.
Hopper, L. M., S.A. Price, H.D. Freeman, S.P. Lambeth, S.J. Schapiro, and R.L. Kendal. 2014. Influence of personality, age, sex, and estrous state on chimpanzee problem-solving success. Animal Cognition 17(4): 835–847. DOI: 10.1007/s10071-013-0715-y.
Keeley, L. H., and N. Toth. 1981. Microwear polishes on early stone tools from Koobi Fora, Kenya. Nature 293: 464–465. DOI: 10.1038/293464a0.
Koops, K., W.C. McGrew, and T. Matsuzawa. 2010. Do chimpanzees (Pan troglodytes) use cleavers and anvils to fracture Treculia africana fruits? Preliminary data on a new form of percussive technology. Primates 51: 175–178. DOI: 10.1007/s10329-009-0178-6.
Koops, K., E. Visalberghi, and C.P. van Schaik. 2014. The ecology of primate material culture. Biology Letters 10: 20140508.
Kühl, H. S., A.K. Kalan, M. Arandjelovic, F. Aubert, L. D’Auvergne, A. Goedmakers, S. Jones, L. Kehoe, S. Regnaut, A. Tickle, E. Ton, et al. 2016. Chimpanzee accumulative stone throwing. Scientific Reports 6(1): 22219. DOI: 10.1038/srep22219.
Luncz, L. V., R. Mundry, and C. Boesch. 2012. Evidence for cultural differences between neighboring chimpanzee communities. Current Biology 22: 922–926.
Marchant, L. F., and W.C. McGrew. 2005. Percussive technology: Chimpanzee baobab smashing and the evolutionary modelling of hominin knapping. In Stone knapping: The necessary conditions for a uniquely hominin behaviour, ed. by V. Roux, and B. Bril. McDonald Institute for Archaeological Research.
Marlowe, F.W. 2005. Hunter-gatherers and human evolution. Evolutionary Anthropology: Issues, News, and Reviews 14(2): 54–67. DOI: 10.1002/evan.20046.
McGrew, W. C. 2010. Chimpanzee technology. Science 328: 579–580.
Moore, M.W. 2020. Hominin Stone Flaking and the Emergence of ‘Top-down’ Design in Human Evolution. Cambridge Archaeological Journal 30: 647–664. DOI: 10.1017/S0959774320000190.
Morgan, T.J., N.T. Uomini, L.E. Rendell, L. Chouinard-Thuly, S.E. Street, H.M. Lewis, C.P. Cross, C. Evans, R. Kearney, I. de la Torre, A. Whiten, and K.N. Laland. 2015. Experimental evidence for the co-evolution of hominin tool-making teaching and language. Nat Commun 6: 6029. DOI: 10.1038/ncomms7029.
Motes-Rodrigo, A., S.P. McPherron, W. Archer, R.A. Hernandez-Aguilar, and C. Tennie. 2022. Experimental investigation of orangutans’ lithic percussive and sharp stone tool behaviours. PLOS ONE 17(2): e0263343. DOI: 10.1371/journal.pone.0263343.
Motes-Rodrigo, A., R. Mundry, J. Call, and C. Tennie. 2021. Evaluating the influence of action- and subject-specific factors on chimpanzee action copying. Royal Society Open Science 8(2): 200228. DOI: 10.1098/rsos.200228.
Mountford, C. P. 1941. An unrecorded method of manufacturing wooden implements by simple stone tools. Transactions of the Royal Society of South Australia 65: 312–317.
Neadle, D., E. Bandini, and C. Tennie. 2020. Testing the individual and social learning abilities of task-naïve captive chimpanzees (Pan troglodytes sp.) in a nut-cracking task. PeerJ 8: e8734.
Nonaka, T., B. Bril, and R. Rein. 2010. How do stone knappers predict and control the outcome of flaking? Implications for understanding early stone tool technology. Journal of Human Evolution 59(2): 155–167. DOI: 10.1016/j.jhevol.2010.04.006.
Panger, M. A., A.S. Brooks, B.G. Richmond, and B. Wood. 2002. Older than the Oldowan? Rethinking the emergence of hominin tool use. Evol. Anthropol. 11: 235–245. DOI: 10.1002/evan.10094.
Pargeter, J., N. Khreisheh, and D. Stout. 2019. Understanding stone tool-making skill acquisition: Experimental methods and evolutionary implications. Journal of Human Evolution 133: 146–166. DOI: 10.1016/j.jhevol.2019.05.010.
Pope, S. M., J.P. Taglialatela, S.A. Skiba, and W.D. Hopkins. 2018. Changes in frontoparietotemporal connectivity following do-as-I-do imitation training in chimpanzees (Pan troglodytes). J. Cogn. Neurosci 30: 421–431.
Potts, R., and P. Shipman. 1981. Cutmarks made by stone tools on bones from Olduvai Gorge, Tanzania. Nature 291: 577–580. DOI: 10.1038/291577a0.
Proffitt, T., L.V. Luncz, T. Falotico, E.B. Ottoni, I. de la Torre, and M. Haslam. 2016. Wild monkeys flake stone tools. Nature 539(7627): 85–88. DOI: 10.1038/nature20112.
Rutz, C., and J.J.H. St Clair. 2012. The evolutionary origins and ecological context of tool use in New Caledonian crows. Behavioural Processes 89(2): 153–165. DOI: 10.1016/j.beproc.2011.11.005.
Savage-Rumbaugh, S., and W.M. Fields. 2006. Rules and tools: Beyond anthropomorphism. In The Oldowan: Case studies into the earliest stone age, ed. by N. Toth, and K. Schick, pp. 223–241. Stone Age Institute Press.
Savage-Rumbaugh, S., K. McDonald, R.A. Sevcik, W.D. Hopkins, and E. Rubert. 1986. Spontaneous Symbol Acquisition and Communicative Use By Pygmy Chimpanzees (Pan paniscus). Journal of Experimental Psychology 115: 211–235.
Schick, K. D., N. Toth, G. Garufi, E.S. Savage-Rumbaugh, D. Rumbaugh, and R. Sevcik. 1999. Continuing investigations into the stone tool-making and tool-using capabilities of a bonobo (Pan paniscus). Journal of Archaeological Science 26: 821–832. DOI: 10.1006/jasc.1998.0350.
Shea, J. J. 2011. Stone tool analysis and human origins research: Some advice from uncle Screwtape. Evolutionary Anthropology: Issues, News, and Reviews 20(2): 48–53. DOI: 10.1002/evan.20290.
Shumaker, R. W., K.R. Walkup, and B.B. Beck. 2011. Animal tool behavior: The use and manufacture of tools by animals. Johns Hopkins University Press.
Snyder, W. D., J.S. Reeves, and C. Tennie. 2022. Early knapping techniques do not necessitate cultural transmission. Science Advances 8(27): eabo2894. DOI: 10.1126/sciadv.abo2894.
Snyder, W. D., J.S. Reeves, and C. Tennie. (in press). Early knapping techniques do not necessitate cultural transmission. Science Advances.
Stout, D., E. Hecht, N. Khreisheh, B. Bradley, and T. Chaminade. 2015. Cognitive Demands of Lower Paleolithic Toolmaking. PLoS One 10: e0121804.
Stout, D., and S. Semaw. 2006. Knapping skill of the earliest stone toolmakers: Insights from the study of modern human novices. In The Oldowan: Case Studies Into the Earliest Stone Age, ed. by K. D. Schick, and N. Toth, pp. 307–320. Stone Age Institute Press.
Tennie, C., D.R. Braun, L.S. Premo, and S.P. McPherron. 2016. The Island Test for Cumulative Culture in the Paleolithic. In The Nature of Culture, ed. by M. N. Heidle, N. J. Conard, and M. Bolus, pp. 121–133. Springer.
Tennie, C., J. Call, and M. Tomasello. 2012. Untrained chimpanzees (Pan troglodytes schweinfurthii) fail to imitate novel actions. PLoS One 7(8): e41548. DOI: 10.1371/journal.pone.0041548.
Tennie, C., L.S. Premo, D.R. Braun, and S.P. McPherron. 2017. Early Stone Tools and Cultural Transmission: Resetting the Null Hypothesis. Current Anthropology 58(5): 652–672. DOI: 10.1086/693846.
Tomasello, M., and J. Call. 2004. The role of humans in the cognitive development of apes revisited. Anim Cogn 7: 213–215. DOI: 10.1007/s10071-004-0227-x.
Tomasello, M., S. Savage-Rumbaugh, and A.C. Kruger. 1993. Imitative Learning of Actions on Objects by Children, Chimpanzees, and Enculturated Chimpanzees. Child Development 64: 1688–1705.
Toth, N. 1982. The Stone Technologies of Early Hominids at Koobi Fora, Kenya: An Experimental Approach [PhD dissertation]. Department of Anthropology, University of California.
Toth, N. 1985. The Oldowan reassessed: A close look at early stone artifacts. Journal of Archaeological Science: Reports 12: 101–120.
Toth, N., K.D. Schick, E.S. Savage-Rumbaugh, R.A. Sevcik, and D.M. Rumbaugh. 1993. Pan the Tool-Maker: Investigations Into the Stone Tool-Making and Tool-Using Capabilities of a Bonobo (Pan paniscus). Journal of Archaeological Science: Reports 20: 81–91.
Toth, N., K. Schick, and S. Semaw. 2006. A comparative study of the stone tool-making skills of Pan, Australopithecus, and Homo sapiens. In The Oldowan: Case studies into the earliest Stone Age, ed. by N. Toth, and K. Schick. Stone Age Institute Press.
Westergaard, G. C., and S.J. Suomi. 1994. A simple stone-tool technology in monkeys. Journal of Human Evolution 27: 399–404. DOI: 10.1006/jhev.1994.1055.
Whiten, A., K. Schick, and N. Toth. 2009. The evolution and cultural transmission of percussive technology: integrating evidence from palaeoanthropology and primatology. Journal of Human Evolution 57: 420–435. DOI: 10.1016/j.jhevol.2008.12.010.
Wright, R. V. S. 1972. Imitative Learning of a Flaked Stone Technology—The Case of an Orangutan. Mankind 8: 296–306. DOI: 10.1111/j.1835-9310.1972.tb00451.x.
Wynn, T., R.A. Hernandez-Aguilar, L.F. Marchant, and W.C. McGrew. 2011. “An ape’s view of the Oldowan” Revisited. Evolutionary Anthropology 20: 181–197. DOI: 0.1002/evan.20323.