Zusammenfassung
Wissenschaftliche Diskussionen über die Entstehung afrikanischer Fundstellen des ‘Early Stone Age’, die Beschaffung tierischer Beute durch Homininen, die Regelmäßigkeit des Fleischkonsums oder die Nutzung zentraler Plätze stützen sich hauptsächlich auf taphonomische Studien einiger weniger archäofaunistischer Vergesellschaftungen, wie FLK Zinj (1,84 Ma [Millionen Jahre] alt), Olduvai-Schlucht Bed I, Tansania). Die DS-Fundstelle liegt auf der gleichen Paläooberfläche wie FLK Zinj und stellt eine ebenso gut erhaltene, vertikal eigenständige Ablagerung dar. Die Fundstelle wurde in den letzten Jahren umfangreich ausgegraben (554 m²) und bietet eine unschätzbare Gelegenheit, wichtige und lange diskutierte Fragen zur Lebensweise der frühen Homininen mit neuen Ansätzen auf der Grundlage robuster und zuverlässiger statistischer Methoden, einschließlich des maschinellen Lernens, anzugehen. Auch taphonomische Studien können von der Einbeziehung der räumlichen Komponente in die Interpretation der Entstehung und Funktionalität einer Fundstelle stark profitieren. Die Ergebnisse der taphonomischen und räumlichen Untersuchung von DS deuten auf die folgenden verhaltensmäßigen Komponenten der Lebensweise von Homininen in Bezug auf ihre Nahrungsbeschaffungsstrategien hin: 1) regelmäßiger primärer Zugang zu Tierbeute, 2) selektiver Erwerb von Tierbeute und 3) kollektiver Verzehr von Fleisch an zentralen Verpflegungsplätzen. Diese Merkmale deuten auch auf Kooperation und gemeinsame Nutzung von Nahrung hin. Die Erkenntnisse aus DS deuten darauf hin, dass der frühe Homo im Vergleich zu heutigen Primaten eine erhebliche Verhaltenskomplexität und eine zusammenhängende Sozialstruktur aufwies. Zwei weitere neu entdeckte, anthropogen beeinflusste Fundorte aus derselben Zeit (PTK und AGS, Bed I Olduvai-Schlucht), die sich derzeit in Analyse befinden, werden bald weiteres Licht auf diese Anpassungen werfen.
Introduction
The origins of human behavior have been associated with fundamental adaptations related to subsistence and the acquisition of food resources by hominins, such as food transport to central provisioning places, food sharing and cooperation (Isaac 1978). These characteristics of human behavior may already be reflected in some East African early sites around 2 Ma (million years) ago (e.g., Isaac 1978, 1983; Bunn 1981; Domínguez-Rodrigo et al. 2007, 2021a; Bunn and Pickering 2010; Domínguez-Rodrigo and Pickering 2017). Archaeological sites where taphonomic analyses have determined a prominent anthropogenic agency in accumulation and modification are a minority in the Oldowan archaeological record (~2.6 – 1.7 Ma); however, they are essential for the reconstruction of early human behavior, and the zooarchaeological and taphonomic studies of the archaeofaunal remains at early sites play a key role in these discussions. After decades of intensive research, taphonomists have gained confidence that at least some of the African early Pleistocene assemblages were created by hominins, who transported and butchered carcasses they obtained through primary access (Bunn 1981; Domínguez-Rodrigo et al. 2007, 2021a; Bunn and Pickering 2010; Ferraro et al. 2013; Domínguez-Rodrigo and Pickering 2017; Parkinson 2018).
However, the paucity of the archaeological record from this period has left many unanswered questions or precluded wider behavioral interpretations. Although the number of Oldowan sites has increased notably in the past decades, well-preserved, vertically discrete, extensively excavated hominin-made assemblages from around 2 Ma ago are scarce. Until recently, only FLK Zinj from Bed I in Olduvai Gorge (Tanzania) fit this description (Bunn and Kroll 1986; Domínguez-Rodrigo et al. 2007). Other well-known and prolific regions preserving early assemblages that have provided data for numerous studies, like the ones recovered at Kanjera South or at Koobi Fora (Kenya), dated to 2.0 – 1.6 Ma, have also been interpreted as the result of hominins having primary access to the accumulated and butchered carcasses (Bunn et al. 1980; Plummer et al. 2009; Plummer and Bishop 2016; Oliver et al. 2019; Parkinson et al. 2022). Kanjera South, however, is a time-averaged deposit that could represent multiple depositional processes and agencies (Ferraro et al. 2013). Recent analyses of three archaeofaunal assemblages from Okote Member at Koobi Fora, FwJj14A, FwJj14B, and GaJi14 (~1.5 Ma) also show that H. erectus had access to well-fleshed carcasses (Pobiner et al. 2008), although, surprisingly, stone tools have not been found in association with faunal remains at these sites.
The recent discovery of DS (David’s Site, 1.84 Ma) in Olduvai Gorge and two additional anthropogenic accumulations on the same paleosurface as FLK Zinj (PTK and AGS) that are well-preserved, vertically-discrete and extensive deposits, with clear functional association between stone tools and faunal remains, represents a new and invaluable opportunity to address key and long debated issues regarding early hominin lifestyles: the identification of the main agents of site formation, the hunting or scavenging behavior of hominins, the socioeconomic function of sites, and the behavioral complexity of hominins relative to extant primates.
Apart from the availability of high-integrity and high-resolution archaeological data, the possibility of addressing these issues also rests upon the articulation and structuring of hypotheses about human behavior. Since behavioral features are often functionally interrelated and dependent on each other, they can be contemplated within a systemic view of human behavior (Brooks et al. 1991). This also means that hypotheses about human behavior would also be interconnected. The support of one particular hypothesis thus enhances the heuristic value of the associated hypotheses (Popper 1972; Niiniluoto 1987; Bunge 1998; Domínguez-Rodrigo 2012). This view of behavior increases the amount of knowledge that can be empirically contrasted and helps us ask wider questions about the evolution of human behavior. For example, increased meat-eating is expected to have had physical, cognitive and socioecological consequences and affected broader patterns of adaptation (Isaac 1978, 1983; Bunn 1981; Fowley 1996; Domínguez-Rodrigo et al. 2007). Moreover, meat-eating is not independent from the fact that it occurred on specific locations where materials accumulated in very high densities, and food sharing requires food to be brought back to a protective spot to be distributed. The intertwined hypotheses contrasted here are framed on the theory that ‘human behavior emerged when subsistence was based on a cooperative and solidarious social organization’ (Domínguez-Rodrigo 2012).
In this paper we therefore aim to answer the following questions based on the available taphonomic evidence from DS: Were hominins having regular primary access to animal carcasses? Were hominins selectively hunting small and medium-sized ungulates? Did collective animal carcass consumption take place in central-provisioning places? And did hominins already engage in cooperative and food sharing behaviors?
Today, after a period of considerable development including experimental and actualistic work, archaeological taphonomy counts with a substantial number of approaches and methods that allow the scientific contrasting of hypotheses and assumptions (Lyman 1994; Domínguez-Rodrigo et al. 2011, 2014). This does not mean, however, that each of the taphonomic aspects usually explored in archaeofaunal studies (site formation, skeletal part profiles, bone breakage patterns, bone surface modifications and prey mortality profiles) do not have limitations. In fact, taphonomic methods still face a number of challenges, mainly dealing with small sample sizes, subjectivity when interpreting certain bone surface marks or lack of frames of reference that lead to scenarios of equifinality. Some of these limitations can be mitigated through the use of more robust statistical methods including the combination of variables designed to overcome equifinality and the use of computer vision to identify bone surface modifications (e.g., Domínguez-Rodrigo and Yravedra 2009; Domínguez-Rodrigo and Baquedano 2018; Domínguez-Rodrigo 2019; Jiménez-García et al. 2020).
By means of several examples of the analyses included in the taphonomic study of DS (Cobo-Sánchez 2020) that contain the most relevant evidence regarding hominin foraging behavior, we intend to shed light on the questions outlined above. The complete taphonomic study of DS, however, includes an analysis of site integrity, comprising the examination of bone orientation patterns, specimen size distribution and bone shape and composition. This analysis is used to test the hypotheses that the site is autochthonous, that the distortion of the assemblage through water currents can be excluded, and thereby that the spatial properties of the assemblage have remained intact (Domínguez-Rodrigo et al. 2019a). Skeletal part abundances were analyzed with relation to food utility and return rates. Long limb bone representation was compared to meaty long bone element ratios at other Paleolithic sites in an effort to address site functionality and hominin carcass transport strategies. Bone breakage patterns were approached on the one hand following traditional methods, including a number of indices, ratios, comparison of means and confidence intervals of breakage planes, as well as correspondence analyses of notch type distributions, and on the other hand using machine learning analyses combining several variables (Moclán et al. 2019). Similarly, bone surface modification analyses also included several different approaches that ranged from univariate comparisons of cut mark, percussion mark and tooth mark frequencies with experimental data and with data from other sites to machine learning analyses combining several variables. The same datasets were used in the different approaches in order to show how the latter really contribute to eliminate equifinality in every analysis.
Most of the available experimental data modeling different types of access to carcasses by hominins and carnivores was incorporated. Additionally, the anatomical location of cut marks was analyzed in addition to their frequencies. The taphotype approach, which is very useful to classify assemblages as hominin- or carnivore-made according to the predominant patterns of long bone portion deletion and modification (Domínguez-Rodrigo et al. 2015b), has also been applied. Finally, bovid mortality profiles were analyzed using multivariate statistics in addition to triangular graphs, hoping to overcome the limitations of equifinality sometimes yielded by the latter, and to gain more insight into the differences between carnivores and humans in prey distribution by age.
The taphonomic study of the archaeofauna at DS also included comparisons with other Paleolithic sites, especially with other Bed I sites, but also with sites from more recent periods and other geographical locations where anthropogenic accumulations were purportedly created by hominins having primary access to carcasses mainly through hunting. In all of the presented analyses the data from FLK Zinj was included in combination with data from DS with the intention to find similarities and differences between both pene-contemporaneous sites, and to try to assess hominin behavioral variability during Olduvai Bed I times. The following section introduces the DS site and the next sections outline how the three main analyses and results support each of the aforementioned hypotheses, and draws an overall picture of hominin lifeways 1.84 Ma ago in Olduvai Gorge.
The DS Site
Olduvai Gorge is located in northern Tanzania, southeast of the Serengeti Plains (Fig. 1A). The oldest sediments it contains are around 2 Ma old. Over the past 200 ka (thousand years) fluvial erosion has carved through the different geological layers creating the gorge, which splits into two branches, the main and the secondary gorge. Hay (1976) defined several geological units within the gorge that are still the foundation for geological studies at Olduvai today: Beds I, II, III, IV, the Masek, Ndutu and Naisiusiu Beds. DS lies in Bed I, which spans from approximately 1.98 Ma to 1.75 Ma, and is formed by the alternating deposition of clayey facies associated to lake sediments and silty layers on the one hand and volcanic tuffs (1A – 1F) that have been securely dated on the other hand (Walter et al. 1991, 1992; Manega 1993; Blumenschine et al. 2016).
Several important sites are known from the layers occurring between these tuffs, like FLKN, FLKNN and DK, but the four fully anthropogenic sites discovered in Bed I (FLK Zinj, PTK, DS, and AGS) all stem from the paleosol underlying Tuff 1C, which contains a clay stratum (<20 cm; Levels 22A and 22B) that can be traced laterally on a significant portion of the gorge at the junction and on both ends of its trajectory in areas that lay close to an ancient lake (Fig. 1B, 1C; Fig. 2).

DS (1.84 Ma) is located at the junction between the main and secondary branches of Olduvai Gorge in northern Tanzania (2°59’33″S; 35°21’08″E). It was discovered a few hundred meters away from FLK Zinj, in an area covered by a dirt road. Excavations started immediately after its discovery revealing dense clusters of fossils and stone tools in situ, and went on for five field seasons (Fig. 1B). From 2014 to 2018, intensive excavations exposed an area of 554 m². DS is therefore larger than the pene-contemporaneous FLK Zinj and FLK NN 1 sites and constitutes the biggest open window to the African Early Pleistocene to date. Over the course of these five fieldwork seasons, more than 15,000 fossil remains and stone tools (including sieve finds) were recovered from Level 22B. Level 22A contained archaeological remains as well, yet in much lower densities. Excavations proceeded quickly during the first two field seasons, because the archaeological deposit was found at just a few centimeters below the surface. Excavations of DS were initially conducted on the basalt platform and at the outcrop on both sides of the road. The arc-shaped excavation is caused by an erosive gap in between both areas.
DS stands out not only for the density of materials, but also for their spatial extension. By the end of the 2015 field season, excavations had already exposed a large area. At this point, a spatial statistical prediction using different regression models and simulations was carried out in order to detect the potentially densest portions of the unexcavated areas of DS (Domínguez-Rodrigo et al. 2017). The high correspondence between the results obtained from these predictions and subsequent excavations at the site showed the great potential of the application of spatial statistical techniques in archaeological research.

At the end of the 2016 field season, most of the paleosurface (468 m²) had been exposed and abundant fossil bones and lithic remains had been retrieved from both archaeological levels. In addition, a series of sediment samples were taken from some of the trenches for chemical biomarker analyses. The trenches excavated in 2017 towards the south of the site area yielded somewhat lower fossil densities than those excavated in previous field seasons, suggesting that the limits of the site towards that direction could be close. The trend observed for level 22A was the opposite, however.
In 2018, several additional trenches were opened with the intention of demarcating the site boundaries. Indeed, these excavated trenches yielded lower densities of archaeological materials, establishing site limits and excavations were finalized. It was determined that some of the edges of the site were delimited by erosion. This was especially true of the central area where deposition and erosion of the Ndutu unconformable sediments exposed the underlying clay to modern erosion. In addition, the southern edge of the excavation began yielding lower densities of materials due to a change in facies from clayey to silty sediments, which probably indicated a change in the paleolandscape. The remaining areas are limited by the lava flows of the bottom of Bed I that were already part of the paleolandscape when hominins created the site (Uribelarrea et al. 2014) (Fig. 3a).

Preservation of the fossil bones at DS is comparable to that at FLK Zinj. This suggests that post-depositional processes had a marginal effect on the assemblage. The numerous rib and vertebrae remains recovered support this inference, because these usually disappear due to the action of carnivores or hydraulic processes (Domínguez-Rodrigo et al. 2018). The site seems to have formed in a short period of time, less than two or three years (Domínguez-Rodrigo et al. 2019b), and the ecological conditions at the site can be reconstructed with very high resolution. Thus, many taphonomic, behavioral, and ecological relationships can be explored spatially.
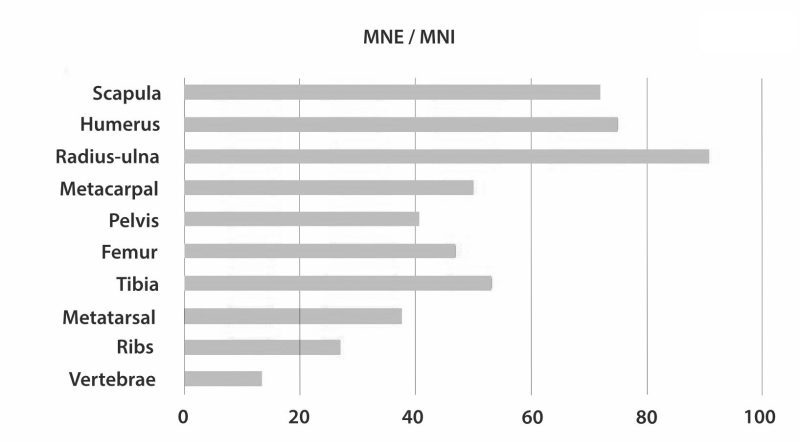
Level 22B from DS contains around 85% of the faunal remains recovered from the site. This level has yielded 3458 bone fragments and 1182 lithics (if counting the fraction longer than 20 mm). The average fossil density in this archaeological level is about eight pieces per square meter, but archaeological materials cluster around three areas of very high fossil density (around 70 remains per square meter) (Fig. 3b). The opposite occurs at PTK, FLK Zinj and AGS where the bulk of the archaeofaunal assemblage and the associated lithic tools come from the uppermost section of the Zinj clay (i.e., 22A) (Leakey 1971; Stancampiano et al. 2022; Organista et al. in press). The zooarchaeological and taphonomic analysis summarized here focused on the ungulate remains from Level 22B at DS. The taphonomic study focused almost exclusively on the ungulate remains, because they represent the bulk of the bone assemblage (98.8%) and also reflect hominin activity most directly, which was the primary aim of the study. A comprehensive analysis of the remaining faunal sample, which mainly includes birds and small carnivores, could provide relevant information about the ecological conditions at the site. Level 22A has also provided evidence of hominin activity at the site yet, it seems, not as conspicuous as Level 22B.
Regular primary access to animal carcasses
Addressing whether hominins had early and primary access to carcasses is necessary to justify food surplus and intentional food sharing. If abundant meat and butchery and primary access to carcasses is confirmed then active foraging strategies, food transport, food sharing and cooperation are more likely, since primary access is a prerequisite for these behaviors.
Primary access to carcasses at DS would be supported by high frequencies of cut marked and percussion marked bone specimens that coincide with the corresponding experimental scenarios (e.g., Domínguez-Rodrigo 1997; Domínguez-Rodrigo and Barba 2005; Galán et al. 2009), by the location of cut marks on hot zones in long bones evidencing filleting (Domínguez-Rodrigo et al. 2007), and on ribs and/or vertebrae evidencing evisceration (Binford 1981; Bunn 1982). We would also expect a high proportion of dynamic long bone breakage (including a high number of impact flakes and type A notches) (e.g., Moclán and Domínguez-Rodrigo 2018), and lack of typical felid damage patterns on the bones (including taphotypes typically associated with felids). In addition, we should find taxonomic diversity that would contrast with a highly specialized felid predatory range (Domínguez-Rodrigo 2012).
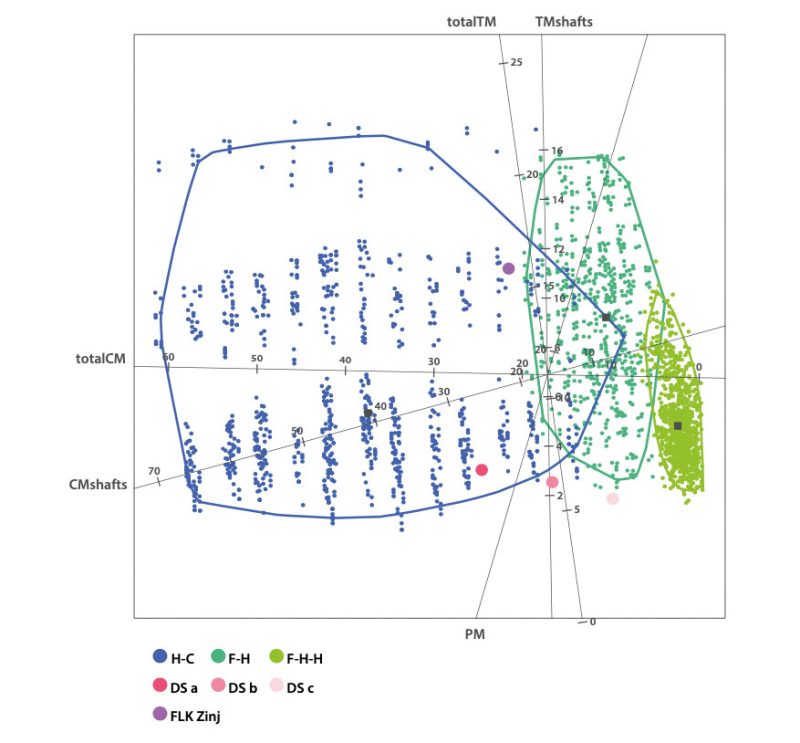
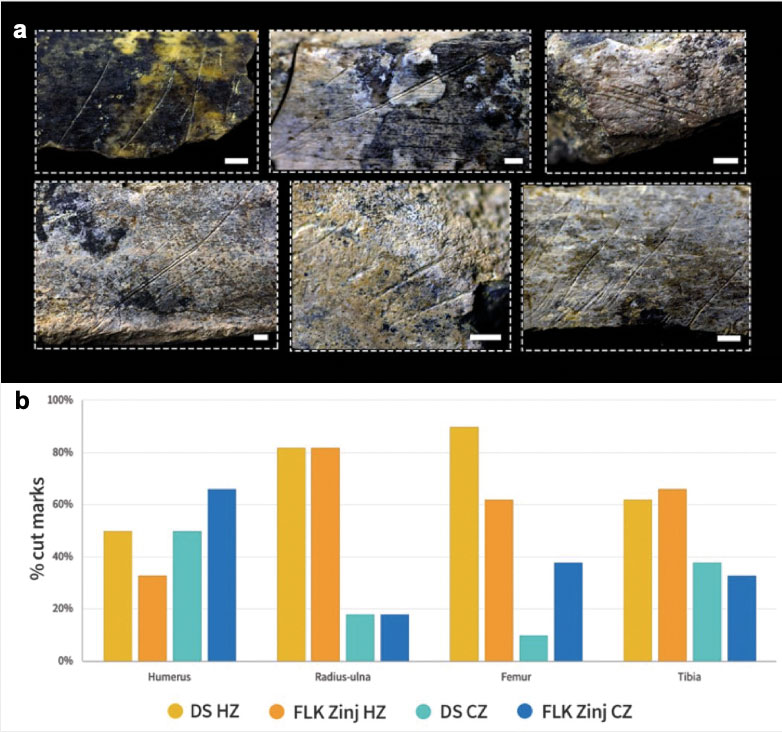
Systematic evidence of exclusive hominin flesh exploitation and evidence of the complete butchering process of several carcasses, from evisceration to the access to the marrow content of bones, supports that meat was consumed abundantly at DS. Moreover, the accumulation on the same spot of several animals spanning carcass sizes from 1 to 3-4 (following Brain 1981), especially of animals that weigh >100 kg, that show evidence of having been defleshed entirely or primarily by hominins, suggests that meat was sufficiently abundant to be shared collectively. Both small and medium-sized carcasses present high frequencies of cut marked and percussion marked bones that coincide with hominin-to-carnivore scenarios. As an example, Figure 4 and Tables 1 and 2 show the results of the classifications of the DS and FLK Zinj bone surface mark samples using a multiple discriminant analysis with a canonical variate approach on a bootstrapped sample of the experimental assemblages of the Hominin-Carnivore (dark blue alpha bag), Felid-Hominin (dark green alpha bag), Felid-Hominin-Hyenid (light green alpha bag) models. Table 3 shows the classification results of several statistical machine learning algorithms into primary or secondary access of DS. All methods yield 100% accuracy. Classifications were nearly always higher than 95% and overall, DS is classified as primary access more than 80% of the times. Moreover, cut mark percentages coincide with those documented at several Upper Paleolithic sites as well as those reported for FLK Zinj (Cobo-Sánchez 2020). Cut marks have been found on all skeletal parts, including ribs (indicating that animals were eviscerated by hominins), vertebrae and skulls. Additionally, most cut marks appear on midshafts of meaty long bones, which suggests not much disarticulation was carried out (Fig. 5a). In fact, the systematic presence of cut marks on hot zones reveals that hominins were accessing fleshed carcasses (Fig. 5b). Lower cut mark percentages are documented on LLB than on ULB and ILB. Several fragments present both cut marks and percussion marks. Only 5% of the long bone MNE were complete, and fragmentation ratios are high, suggesting that marrow was intensively exploited. Most bone breakage is also attributed to hominins by machine learning algorithms (Cobo-Sánchez 2020).

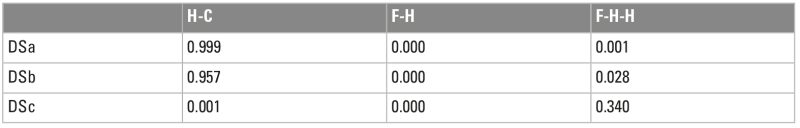
With regard to carnivore damage specifically, the low tooth mark percentages documented on different portions of the bones, also mostly coincide with hominin-to-carnivore models. The presence of felid modification patterns on the carcasses would be indicative of scavenging. Typical modification patterns in felid-consumed carcasses include deep pits on the mesio-lateral sides of the distal femoral troclea, damage on the caudal medial epicondyle of humeri, furrowing of the olecranon of the ulna, on the tibial crest, the pelvic crest, the scapula blade and on the vertebral apophyses, and pits or punctures on vertebral bodies (Gidna et al. 2015; Domínguez-Rodrigo et al. 2021b). DS lacks these types of modifications. The lack of typical felid damage on bones (taphotypes) associated to felids indicate that hominins were not confronting felids for their prey. Only one specimen shows modifications attributable to a felid, as well as cut marks (Domínguez-Rodrigo et al. 2021a). The remaining tooth marks on DS bone surfaces have all been attributed to hyenas by computer vision through deep learning methods (Cobo-Sánchez et al. 2022). In sum, univariate, multivariate and machine learning analyses of bone surface modifications yield consistent and uniform results pointing overwhelmingly to primary access to carcasses by hominins.

Selective animal carcass acquisition
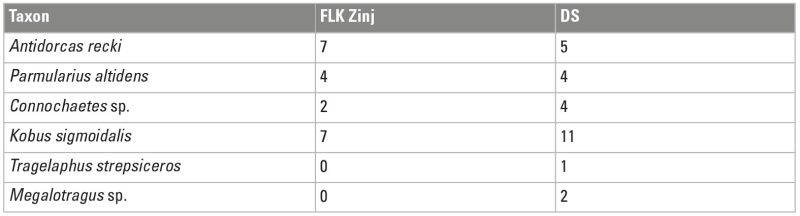
Bovid representation at DS is very similar to that in FLK Zinj. With some exceptions, almost the same taxa are represented at both sites in similar proportions (Table 4). This shows that hominins focused on a range of carcass sizes from 1 to 3-4 and probably targeted certain species and age classes. There are almost 30 bovids represented at DS that reach size 3-4 in their adult stage, as well as five small (size 1-2) bovids and at least five large (size 5) animals. Skeletal part profiles are relatively unbiased, especially those of small carcasses. Small carcass transport seems to have followed an unconstrained strategy; they entered the site complete and were entirely processed. This might be suggestive of short-distance transport from the kill site. In medium-sized carcasses, the appendicular skeleton presents a bias that is not explained in terms of food utility or density-mediated attrition, but seems to reflect particular transport decisions, as explained below (Fig. 6; Cobo-Sánchez 2020).
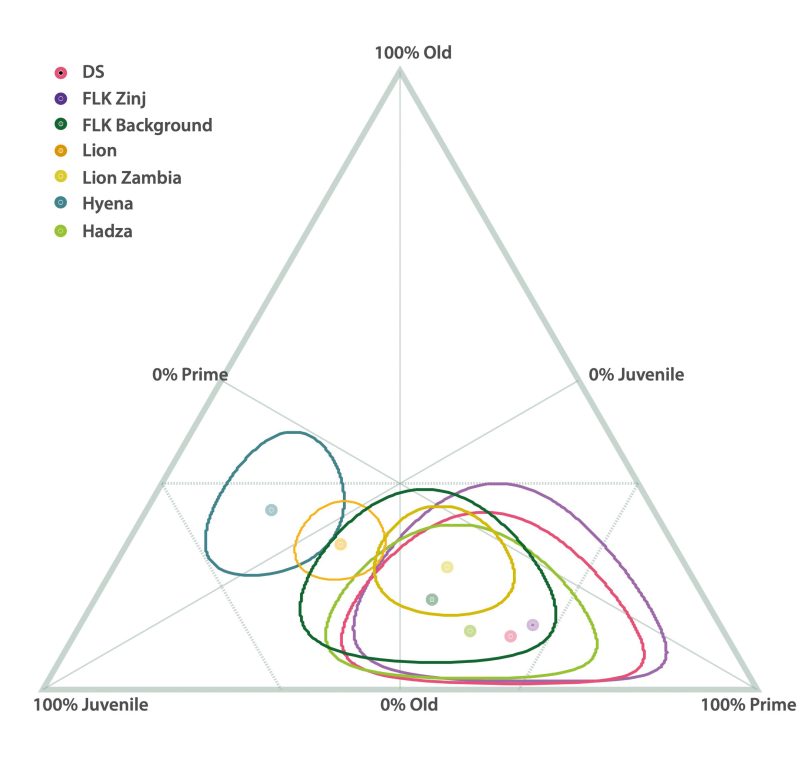
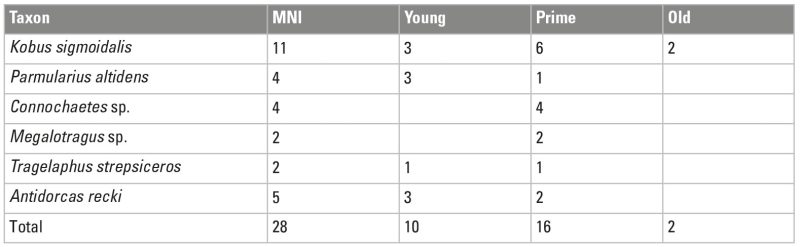
Regular primary access to carcasses as evidenced at DS is necessarily linked to hunting as the main foraging strategy used by hominins to accumulate carcasses at DS. Hunting would be supported by bovid age profiles that differ from the preys of carnivores and/or show similarities to prime adult-dominated profiles typical in many sites of more recent periods of human evolution and age profiles generated by modern hunter-gatherers (e.g. Bunn and Pickering 2010). Hunting would also be supported if the taphotypes did not match those created by felids. It could also be supported potentially by impact marks from sharp or pointed objects on the bones (Gaudzinski-Windheuser et al. 2018). Indeed, several taphonomic results support this foraging strategy. As explained above, taphotypes at DS do not coincide with those typical for felids. Most overlap in the documented mortality profiles of medium-sized bovid carcasses from DS with other assemblages occurs with the Hadza/Kua kills when using the triangle graphs (Fig. 7). Apart from the fact that bovid age profiles at DS do not coincide with age profiles usually created by other carnivores, the bovid accumulations at DS (and FLK Zinj) are both dominated by prime adults (60-70%), something common in many other archaeological sites that are anthropogenic, and which is consistent with ambush hunting (Table 5; Fig. 7).
Large amounts of meat could only have been available regularly for hominins if they acquired carcasses either through hunting and/or through confrontational scavenging, because in savanna biomes only marginal carcass resources are available for scavenging, and only for short periods of time during the end of the dry season (e.g., Blumenschine 1986). Regular confrontational scavenging could therefore also yield relatively fleshed carcasses. However, this foraging strategy should be supported by age profiles similar to the prey targeted by felids, a predominance of typical felid modification patterns (Domínguez-Rodrigo et al. 2015b), and an anatomical distribution and frequency of cut marks and tooth marks that correspond with a felid to hominin to hyenid scenario, none of which characterize the DS archaeofaunal assemblage. As explained above, there is only one documented case of felid damage in the complete assemblage that suggests confrontational scavenging (Domínguez-Rodrigo et al. 2021b). There is, however, a bias in the representation of front and hind limbs consisting in a lower representation of the highest utility parts (hind limbs) that could indicate that some medium-sized carcasses where acquired from confrontational scavenging from felids, stealing carcasses from felid kills that may have lacked viscera and hindlimbs (Fig. 6). Yet, the bias in the representation of front and hind limbs is not necessarily explained by confrontational scavenging. It could also be related to transport distance or simply to variability in hominin transport decisions, and does not exclude hunting.
This means that the detected evidence that could be pointing to confrontational scavenging is inconsistent and ambiguous. For instance, the uneven representation of front and hindlimbs in medium-sized carcasses could mean that hominins butchered and discarded hindlimbs at the kill site, since they have higher return rates and are more efficiently processed, as a strategy to maximize their individual energetic gain and minimize transport load. This behavior would also render the documented skeletal profiles at DS. Regardless, this only affects a small portion of the assemblage; carcasses seem to have been predominantly acquired complete by hominins.
When using the triangle graph, mortality profiles from DS and FLK Zinj overlap partly with one of the age profile samples created by lions and with the FLK background sample. According to this graph, both felids and humans can create very similar mortality patterns that resemble the structure of a living population and show a predominance of prime adults. The mortality profiles from Alcelaphini at DS and one of the lion samples match (Cobo-Sánchez 2020). There is a possibility that alcelaphines from DS were acquired through confrontational scavenging from lions, also given the high variability in age mortality profiles generated by lions (and humans). Yet, as mentioned above, the taphotypes from DS do not overlap with those typical for felids in any case, and overall, the bovid age profiles documented at the site are more similar to mortality profiles created by humans than by felids. Confrontational scavenging could therefore have been part of the hominin behavioral repertoire but, according to the DS data, not the predominant one.
Collective animal carcass consumption in central-provisioning places
A central place has been defined as a locus used repeatedly, to which resources are transported and processed, acting as the focal point of group fission/fusion (Marlowe 2010). The use of the location as a central place would be supported by the presence of local and non-local taxa and a significantly higher bone density at the site than in the surrounding landscape. Additionally, bone clusters should be qualitatively different from those at non-anthropogenic sites, and the accumulation should involve multiple individuals. Presumably, skeletal part profiles would be relatively unbiased. The environmental context of the location would be characterized by low trophic dynamics. A central place would also be characterized by food surplus that would enhance food sharing and evidence of other activities not related to animal carcass consumption. The depositional time should be short (no more than one or two years).
At DS, the high bone density inside the site contrasts significantly with the surrounding landscape, and the assemblage is formed of multiple butchered carcasses belonging to both autochthonous and allochthonous taxa. Taphonomic evidence regarding the site’s integrity excludes the possibility of the accumulation being a death site or the result of natural processes. According to the site formation analysis, the DS archaeofaunal accumulation is autochthonous and undisturbed. Bones show no preferential orientation, and bone shape and composition representation is similar to that of undisturbed assemblages. There is a high degree of fragmentation of the assemblage as opposed to the high percentages of complete bones found in natural assemblages (Cobo-Sánchez 2020). Additionally, the bone clusters are also spatially different from those at non-anthropogenic sites, in essence because they are associated with lithics clusters. The clusters of stone tool debris suggest that aside from butchering, knapping activities took place at DS.
The transport of complete or partial fleshed sections of animals larger than 100 kg (including access to it and initial butchery to be transported) requires the joint participation of several individuals according to carcass size. Moreover, the collective transport of lithic raw material is also inferred, because the amount of lithic artifacts at the site exceeds the physical capability of having been accumulated by one single individual, provided that the accumulation took place mostly in one occupational episode and not across a diachronic sequence of various occupational episodes, as is the case of DS. Abundant lithic raw material exceeding the transport capacity of one individual also refers to anticipation of future needs as reflected in raw material procurement and exploitation and is linked to the reduction sequence concept (Diez-Martín et al. 2021, 2022).
There are abundant axial remains that even appear spatially distributed in six to eight clusters and that show that several carcasses were complete when they were introduced into the site and consumed collectively in reduced areas. The spatial distribution of all butchering-related activities also shows that carcasses were completely butchered in the same spots (Cobo-Sánchez 2020).
The environmental context of the location is characterized by a low to moderate competition and trophic dynamics, as indicated by low to moderate degree of ravaging at the site, and the depositional time does not exceed two or three years, as evidenced by the subaerial weathering stages on the bone surfaces. All these arguments support that DS was used as a central provisioning place for food processing and consumption and stone tool manufacture.
The redundancy in the aforementioned butchery patterns and the representation of resources that clearly exceed the needs of one individual, especially if occurring on the same spot repeatedly, suggests that the purpose of such behavior and use of a central place could have been food sharing.
Discussion
Some authors have argued that hominin carnivory and hominin predatory behaviors evolved gradually from passive scavenging strategies for the obtainment of flesh scraps from felid kills by Oldowan hominins toward consistent early access to carcasses during the Acheulean (e.g., Cachel and Harris 1998; Holliday 2012; Pobiner 2016). Yet, the results of the taphonomic analysis of DS site show that hominins were successful and regular hunters at least 2 Ma ago, and small and medium-sized carcass foraging seem to have emerged as part of a set including other behaviors, like food sharing and cooperation (Pickering and Domínguez-Rodrigo 2010; Domínguez-Rodrigo and Pickering 2017). Interestingly, whenever a strong anthropogenic input is inferred in early Pleistocene archaeofaunal assemblages, the taphonomic evidence points every time to early access to carcasses by hominins, even in older periods in which this evidence in the archaeological record is even more scarce. For example, evisceration marks have been identified at Gona (Ethiopia) at 2.6 Ma (Domínguez-Rodrigo et al. 2005), and at Ain Boucherit (Algeria), which is dated to 2.4 Ma (Sahnouni et al. 2018). It is true that the evidence is not sufficient from 2.6 to 2 Ma to make inferences beyond tentative early access to carcasses, but the subsequent concentrations of lithic artifacts and fossil bones dating from 2.0 Ma and 1.8 Ma like DS and FLK Zinj, which also coincide with the appearance of Homo ergaster (Domínguez-Rodrigo et al. 2015a; Villmoare et al. 2015), allow us to test more assumptions and reconstruct a clearer picture of hominin foraging strategies.
At DS, the results from the bovid age profile analysis support hunting, more specifically (but not exclusively), ambush hunting, mainly because the documented mortality profiles fall inside the range of the age structure of a living population, with a predominance of prime adults, which constitute around 60 to 70% of the medium-sized bovid sample. When it appears in the archaeological and ethnographic record, this pattern is usually associated with selective ambush hunting (e.g., Stiner 1990; Bunn and Pickering 2010). Exactly the same tendency is observed at FLK Zinj for both carcass sizes, especially when using four age classes instead of three. The tendency to hunt high-ranked prey at these early sites also supports that hominins were already efficient hunters, as opposed to the view that their hunting capabilities evolved gradually over the Middle to Upper Pleistocene. Incidentally, hunting scenarios are accepted for several other Lower Paleolithic sites in Eurasia based on similar evidence. Other significant assemblages that are interpreted as the result of hunting are TD6-2 at Gran Dolina in Atapuerca (~0.80 Ma; Saladié et al. 2011, 2014) or the assemblage of fallow deer in Gesher Benot Ya’aqov in Israel (~0.75 Ma; Rabinovich et al. 2008). At TD6-2 all age groups are represented, but the profile is dominated by immature and adult individuals (Saladié et al. 2011, 2014). Gesher Benot Ya’aqov constitutes an assemblage dominated by a single taxon (Dama dama) where adult individuals are most abundant (Rabinovich et al. 2008). At TD10.1 at Gran Dolina in Atapuerca (dated to around ~350 ka), prime-adult individuals dominate the deer mortality profile also. This site represents a long-term residential base camp formed probably by very intensive occupations. This preference on high-ranked prey is similar to that documented in other Middle and Late Paleolithic sites across Europe and the Levant (Rodríguez-Hidalgo et al. 2015, 2017) and coincides with the prey selection profile that is interpreted as unique for humans (Stiner 1990). If we accept the age mortality profiles from these sites and more recent periods as evidence of hominin predatory behavior, the same should apply for earlier anthropogenic sites, especially if they appear coupled with taphonomic evidence for early access, as is the case of DS.
Age profiles are not the only line of evidence with similar outcomes at DS and at sites from other more recent periods. In fact, one of the most important results obtained in the present study is precisely that several aspects of the taphonomic pattern yielded by the faunal assemblage at DS, in particular the frequencies and location of cut marks, do not differ from those obtained from sites from substantially later periods, where hunting is undisputed. The previously mentioned assemblages are also a good example thereof, because they have been studied thoroughly from a taphonomic perspective. Similar as to what can be observed at DS, in these assemblages there is evidence for the performance by hominins of all butchering-related activities, which again establishes that hominins had early access to abundant meat resources. For example, skinning is evidenced by cut marks on the crania, the metapodials and the phalanges. Detachment of the crania from the postcranial skeleton by cut marks on atlases and cut marks (and in some cases also hack marks) on epiphyses of long bones reveal disarticulation activities. Evisceration is demonstrated by abundant cut marks on the ventral sides of ribs and some vertebrae, and intensive defleshing is evidenced by high percentages of cut (slicing) marks on hot zones of meaty long bones. This demonstrates that carcasses were completely and efficiently processed regularly at these sites, following the same butchery patterns that are also typical in later periods of human evolution (e.g., Rabinovich et al. 2008).
The bovid age profiles and the taphonomic and zooarchaeological similarities between DS and other early Pleistocene sites where hunting is accepted are the most important arguments stemming from this study in favor of an early human predatory behavior. However, the fact that the mortality profiles from DS do not consistently overlap with those typical for felid kills is equally important, although occasional overlap is documented with the age profiles created by lions when using three age classes or when using five age classes and considering only the Alcelaphini subsample. However, this match must be taken cautiously, first because lions produce very variable bovid age profiles depending on certain environmental aspects and seasonality, and secondly, because in the case of DS, these similar profiles do not appear in conjunction with typical felid damage on the bones. The almost complete absence of this type of damage patterns in the faunal remains of DS argues against scavenging (passive or confrontational). If most carcasses accumulated at DS were the result of aggressive scavenging from felids, these bone modfication patterns should be more commonly documented.
Evidence from other Bed I sites like FLKN 1-6, FLK 10-15, DK and FLKNN, where hominins do not seem to have taken advantage of large amounts of complete bones from felid accumulated remains, should not be disregarded. At these sites, complete bones were left unprocessed by hominins. For example, at FLKN 1-6 hominin activity is documented by the presence of lithics in every level of the long sequence spanning hundreds or thousands of years, yet they do not seem to have engaged in exploiting the remains of felid kills. This suggests that scavenging from felids was not a common activity, although the possibility that occasional scavenging may have happened should not be excluded. Even if the mentioned taphonomic signals were reflecting power scavenging, the patterns are not consistent. The taphonomic signal should be more accentuated if the bulk of the assemblage had been acquired through aggressive scavenging. By contrast, the factors pointing to hunting are much more consistent. Thus, it is probable that hunting was the predominant strategy used by hominins to obtain large quantities of meat already by 2–1.8 Ma ago.
Aside from determining the degree of involvement of hominins in carcass acquisition and meat consumption at early sites, the evidence from DS shows that some early sites were created by hominins repeatedly carrying animal carcasses and stones to particular places in the landscape. The concept of ‘central place’ entails that these places also had a social function in addition to being used for specific activities related to subsistence. This means that hominins could have used these places for sharing food and therefore have stayed at these locations for prolonged periods of time, rather than only have used these spots sporadically as refuge places or stone caches when trying to avoid predation risk.
The possibility of using central places repeatedly for hominin activities would be conditioned on the one hand by the amount of food surplus available for hominin consumption, a condition met in the case of DS, and on the other hand by the degree of carnivore predation risk in the surroundings of the site (Blumenschine 1991). Carnivore competition or predation risk can be assessed by means of reconstructing the paleoenvironment in which the site was formed and by establishing the degree of carnivore ravaging at a site, which can be used as a proxy for carnivore competition. The taphonomic analyses of the Bed I sites showed that carnivores were very active in the lacustrine plain of the former Olduvai lake where these sites formed. Some felids, like leopards, Dinofelis, and solitary lions, were accumulating carcasses at certain locations, probably seeking low-competition settings. The predatory guild was very diverse compared to the carnivore species that are found in modern savannas today, which also suggests that competition among carnivores must have been significant (Domínguez-Rodrigo et al. 2007). Sites were probably located in transitional zones between the alluvial plain and the lacustrine floodplain in closed-vegetation habitats (lacustrine forests), which were probably relatively low-competition habitats (Domínguez-Rodrigo et al. 2007). DS and FLK Zinj were located on a topographically higher platform in a wooded environment dominated by palm and acacia trees (Uribelarrea et al. 2014; Arráiz et al. 2017). These wooded environments would have represented the lowest competition areas, which explains why hominins would have selected these loci for their activities. The analysis of several skeletal part ratios to estimate ravaging intensity also suggested that the degree of ravaging and carnivore competition were low to moderate at DS. More intensive ravaging is documented at FLKN and DK, which formed in a more open environment. The same inferences can be drawn from bovid taxa representation at the sites. While more species from mixed habitats such as waterbucks (Kobus) are predominant at FLK Zinj and DS, bovids from more open environments, like Antidorcas and Parmularius are dominant at FLKN (Domínguez-Rodrigo et al. 2007). The few individuals belonging to these taxa represented at DS were probably carried across longer distances and brought into the sites from further than the immediate vicinity of the site.
High degrees of carnivore competition could have had consequences regarding scavenging opportunities for hominins. If carnivores were very active in the Olduvai Bed I environment, the opportunities for passive scavenging would have been very low. In addition, since confrontational scavenging is a riskier activity than hunting, it is possible that carnivore predation could have posed a major threat for hominins, as they do for modern foragers still today (Treves and Naughton-Treves 1999). Yet, the risk of carnivore predation could have likewise promoted higher cooperative behaviors among hominins and increased sociality, as is observed in fact in several extant primate species (Rose and Marshall 1996).
Bone remains and lithic artifacts at PTK and AGS present similar taphonomic signatures and seem to be also spatially distributed in large clusters of similar characteristics as the ones from FLK Zinj and DS, which could suggest that these sites could have been selected by hominins for similar purposes (work in progress). This does not preclude the use of other locations in the Zinj paleolandscape by hominins for other activities. The palimpsests formed at other Bed I sites show that hominins were active in other loci as well. The existence of at least three central places in the Zinj paleolandscape in a relatively small area (around 1000 m²) suggests that hominin groups were very active in the area, just like carnivores. The floodplain, with several (seasonal) fluvial input areas and the nearby location of a freshwater spring could have been an important area of attraction for many species of herbivores, given the presence of accessible water resources.
Conclusions
The taphonomic study of DS supports previous interpretations about hominin behavior drawn from earlier taphonomic analyses of FLK Zinj, as well as the high explanatory power of the applied taphonomic and spatial approaches. In particular, the results show that the faunal assemblage from Level 22B at DS is an autochthonous largely undisturbed assemblage and that complete fleshed small and medium-sized ungulate carcasses were actively transported and introduced into the site by hominins. These had early access to meat resources. The evidence also points to hunting as the main carcass foraging strategy employed by early humans, probably through ambush techniques, and suggests that carnivore activities were limited at the site. DS seems to have been used by hominins as a central place near locations with easy access to herbivores and water sources. At these places, hominins probably congregated on small areas and consumed food collectively. The regular successful acquisition of large carcasses and their transport to central places likely relied on high levels of group cooperation. The evidence from DS indicates that early Homo showed significant behavioral complexity relative to extant primates and a very cohesive social structure.
These interdependent behaviors inferred from DS and FLK Zinj are probably attributable to Homo ergaster or to a similar species. Therefore, the results are also in line with the general agreement among researchers that the appearance of this species marked a significant shift in adaptation from the other hominins. Around 2 Ma ago, the consumption of meat by hominins increased significantly, and a number of socio-reproductive behaviors that enabled and required meat-eating were also adopted. At that time, hominins appear to have already been part of the predatory guild and very successful hunters. The early archaeological record shows that in spite of their limited technology and their smaller brains compared to those of modern humans, early humans were capable of certain cognitively complex behaviors, including anticipating adaptive needs, and were well-adapted to certain forms of hunting and group cooperation, even though their predatory behaviors and social reproductive structures may not have an equivalent in the present.
Acknowledgments
This paper is partially drawn from my unpublished PhD dissertation, “Taphonomic and spatial study of the archaeological site DS from Bed I (Olduvai Gorge, Tanzania).” I am thankful to my supervisors, Prof. Manuel Domínguez-Rodrigo and Prof. Gonzalo Ruiz Zapatero, and my examiners, Dr. Antonio Rodríguez-Hidalgo, Dr. Palmira Saladié, Dr. Enrique Baquedano, Prof. Nicholas Conard, and Prof. Almudena Hernando for their insightful comments during my defense. I am very thankful to the University of Tübingen committee for awarding me the 2022 Tübingen Research Prize for Early Prehistory and Quaternary Ecology. I also thank Banco de Santander and the Complutense University of Madrid for funding my research. My PhD would not have been possible without all the people involved in the excavations at DS and the restoration of the faunal remains. Fieldwork in Olduvai Gorge was possible thanks to funding from the Spanish Ministry of Science, Innovation and Universities (HAR2017-82463-C4-1-P), the Ministry of Culture through the program of Archaeology Abroad, the Palarq Foundation and E2in2. COSTECH (Commission for Science and Technology), the Division of Antiquities, NCAA (Ngorongoro Conservation Area Authority) and TANAPA (Tanzanian National Parks), within the Tanzanian Ministry of Natural Resources and Tourism granted permission to conduct research in Tanzania.
References
Arráiz, H., Barboni, D., Ashley, G. M., Mabulla, A., Baquedano, E., and Domínguez-Rodrigo, M. 2017: The FLK Zinj paleolandscape: Reconstruction of a 1.84 Ma wooded habitat in the FLK Zinj-AMK-PTK-DS archaeological complex, Middle Bed I (Olduvai Gorge, Tanzania). Palaeogeography, Palaeoclimatology, Palaeoecology 488, 9–20.
Binford, L. R. 1981: Human modes of bone modification. Chapter in: L. R. Binford, Bones. Ancient Men and Modern Myths. New York: Academic Press, 87–181.
Blumenschine, R. J. 1986: Carcass consumption sequences and the archaeological distinction of scavenging and hunting. Journal of Human Evolution 15, 639–659.
Blumenschine, R. J. 1991: Hominid carnivory and foraging strategies, and the socio-economic function of early archaeological sites. Philosophical Transactions of the Royal Society London B, Biological Sciences 334, 211–221 (219–221: discussion).
Blumenschine, R. J., Peters, C. R., Masao, F. T., Clarke, R. J., Deino, A. L., Hay, R. L., Swisher, C. C., Stanistreet, I. G., Ashley, G. M., McHenry, L. J., Sikes, N. E., van der Merwe, N. J., Tactikos, J. C., Cushing, A. E., Deocampo, D. M., Njau, J. K., and Ebert, J. I. 2016: Late Pliocene Homo and hominid land use from western Olduvai Gorge, Tanzania. In: R. L. Ciochon and J. G. Fleagle (eds.), The Human Evolution Source Book, Second edition. New York: Routledge, 255–262.
Brain, C. K. 1981: The Hunters or the Hunted? An Introduction to African Cave Taphonomy. Chicago and London: The University of Chicago Press.
Brooks, D. R. and McLennan, D. A. 1991: Phylogeny, Ecology, and Behavior. A Research Program in Comparative Biology. Chicago: University of Chicago Press.
Bunge, M. 1998: Philosophy of Science. From Explanation to Justification. Volume 2, revised edition. Transaction Publishers. London and New York: Routledge.
Bunn, H. T. 1981: Archaeological evidence for meat-eating by Plio-Pleistocene hominids from Koobi Fora and Olduvai Gorge. Nature 291, 574–577.
Bunn, H. T. 1982: Animal Bones and Archeological Inference. Book review of L. R. Binford, Bones. Ancient Men and Modern Myths (1981, New York: Academic Press). Science 215, 494–495.
Bunn, H. T. and Gurtov, A. N. 2014: Prey mortality profiles indicate that Early Pleistocene Homo at Olduvai was an ambush predator. Quaternary International 322-323, 44–53.
Bunn, H. T. and Kroll, E. M. 1986: Systematic Butchery by Plio/Pleistocene Hominids at Olduvai Gorge, Tanzania. With comments by S. H. Ambrose, A. K. Behrensmeyer, L. R. Binford, R. J. Blumenschine, R. G. Klein, H. M. McHenry and C. J. O’Brien, J. J. Wymer, and a reply by H. T. Bunn and E. M. Kroll. Current Anthropology 27, 431–452.
Bunn, H. T. and Pickering, T. R. 2010: Bovid mortality profiles in paleoecological context falsify hypotheses of endurance running–hunting and passive scavenging by early Pleistocene hominins. Quaternary Research 74, 395–404.
Bunn, H., Harris, J. W. K., Isaac, G., Kaufulu, Z., Kroll, E., Schick, K., Toth, N., and Behrensmeyer, A. K. 1980: FxJj50: An early Pleistocene site in northern Kenya. World Archaeology 12, 109–136.
Cobo-Sánchez, L. 2020: Taphonomic and spatial study of the archaeological site DS from Bed I in Olduvai Gorge (Tanzania). Unpublished PhD dissertation. Universidad Complutense, Madrid.
Cobo-Sánchez, L., Pizarro-Monzo, M., Cifuentes-Alcobendas, G., Jiménez García, B., Abellán Beltrán, N., Courtenay, L. A., Mabulla, A., Baquedano, E., and Domínguez-Rodrigo, M. 2022: Computer vision supports primary access to meat by early Homo 1.84 million years ago. PeerJ 10: e14148.
Diez-Martín, F., Cobo-Sánchez, L., Baddeley, A., Uribelarrea, D., Mabulla, A., Baquedano, E., and Domínguez-Rodrigo, M. 2021: Tracing the spatial imprint of Oldowan technological behaviors: A view from DS (Bed I, Olduvai Gorge, Tanzania). PLoS ONE 16(7): e0254603.
Diez-Martín, F., Panera, J., Maíllo-Fernández, J. M., Santonja, M., Sánchez-Yustos, P., Pérez-González, A., Duque, J., Rubio, S., Marín, J., Fraile, C., Mabulla, A., Baquedano, E., and Domínguez-Rodrigo, M. 2022: The evolution of stone tool technology at Olduvai Gorge (Tanzania): Contributions from the Olduvai Paleoanthropology and Paleoecology Project. L’Anthropologie 126: 103000. https://doi.org/10.1016/j.anthro.2022.103000.
Domínguez-Rodrigo, M. 1997: Testing meat-eating in early hominids: an analysis of butchery marks on defleshed carcasses. Human Evolution 12, 169–182.
Domínguez-Rodrigo, M. 2012: Towards a scientific-realistic theory on the origin of human behavior. In: Domínguez-Rodrigo, M. (ed.), Stone Tools and Fossil Bones. Debates in the Archaeology of Human Origins. New York: Cambridge University Press, 11–44.
Domínguez-Rodrigo, M. 2019: Successful classification of experimental bone surface modifications (BSM) through machine learning algorithms: a solution to the controversial use of BSM in paleoanthropology? Archaeological and Anthropological Sciences 11, 2711–2725.
Domínguez-Rodrigo, M. and Baquedano, E. 2018: Distinguishing butchery cut marks from crocodile bite marks through machine learning methods. Scientific Reports 8:5786.
Domínguez-Rodrigo, M. and Barba, R. 2005: A study of cut marks on small-sized carcasses and its application to the study of cut-marked bones from small mammals at the FLK Zinj site. Journal of Taphonomy 3, 121–134.
Domínguez-Rodrigo, M. and Pickering, T. R. 2017: The meat of the matter: an evolutionary perspective on human carnivory. Azania: Archaeological Research in Africa 52, 4–32.
Domínguez-Rodrigo, M. and Yravedra, J. 2009: Why are cut mark frequencies in archaeofaunal assemblages so variable? A multivariate analysis. Journal of Archaeological Science 36, 884–894.
Domínguez-Rodrigo, M., Pickering, T. R., Semaw, S., and Rogers, M. J. 2005: Cutmarked bones from Pliocene archaeological sites at Gona, Afar, Ethiopia: implications for the function of the world’s oldest stone tools. Journal of Human Evolution 48, 109–121.
Domínguez-Rodrigo, M., Barba, R., and Egeland, C. P. 2007: Deconstructing Olduvai: A Taphonomic Study of the Bed I Sites. Vertebrate Paleobiology and Paleoanthropology Series. Dordrecht: Springer.
Domínguez Rodrigo, M., Fernández-López, S., and Alcalá, L. 2011: How can taphonomy be defined in the XXI century? Journal of Taphonomy 9(1), 1–13.
Domínguez-Rodrigo, M., Bunn, H. T., and Yravedra, J. 2014: A critical re-evaluation of bone surface modification models for inferring fossil hominin and carnivore interactions through a multivariate approach: Application to the FLK Zinj archaeofaunal assemblage (Olduvai Gorge, Tanzania). Quaternary International 322-323, 32–43.
Domínguez-Rodrigo, M., Pickering, T. R., Almécija, S., Heaton, J. L., Baquedano, E., Mabulla, A., and Uribelarrea, D. 2015a: Earliest modern human-like hand bone from a new >1.84-million-year-old site at Olduvai in Tanzania. Nature Communications 6:7987.
Domínguez-Rodrigo, M., Yravedra, J., Organista, E., Gidna, A., Fourvel, J.-B., and Baquedano, E. 2015b: A new methodological approach to the taphonomic study of paleontological and archaeological faunal assemblages: a preliminary case study from Olduvai Gorge (Tanzania). Journal of Archaeological Science 59, 35–53.
Domínguez-Rodrigo, M., Cobo-Sánchez, L., Uribelarrea, D., Arriaza, M. C., Yravedra, J., Gidna, A., Organista, E., Sistiaga, A., Martín-Perea, D., Baquedano, E., Aramendi, J., and Mabulla, A. 2017: Spatial simulation and modelling of the early Pleistocene site of DS (Bed I, Olduvai Gorge, Tanzania): a powerful tool for predicting potential archaeological information from unexcavated areas. Boreas 46, 805–815.
Domínguez-Rodrigo, M., Cobo-Sánchez, L., Yravedra, J., Uribelarrea, D., Arriaza, C., Organista, E., and Baquedano, E. 2018: Fluvial spatial taphonomy: a new method for the study of post-depositional processes. Archaeological and Anthropological Sciences 10, 1769–1789.
Domínguez-Rodrigo, M., Baquedano, E., Barba, R., Uribelarrea, D., and Gidna, A. 2019a: The river that never was: Fluvial taphonomy at Olduvai Bed I and II sites and its bearing on early human behavior. Quaternary International 526, 26–38.
Domínguez-Rodrigo, M., Sánchez-Flores, A. J., Baquedano, E., Arriaza, M. C., Aramendi, J., Cobo-Sánchez, L., Organista, E., and Barba, R. 2019b: Constraining time and ecology on the Zinj paleolandscape: Microwear and mesowear analyses of the archaeofaunal remains of FLK Zinj and DS (Bed I), compared to FLK North (Bed I) and BK (Bed II) at Olduvai Gorge (Tanzania). Quaternary International 526, 4–14.
Domínguez-Rodrigo, M., Baquedano, E., Organista, E., Cobo-Sánchez, L., Mabulla, A., Maskara, V., Gidna, A., Pizarro-Monzo, M., Aramendi, J., Galán, A. B., Cifuentes-Alcobendas, G., Vegara-Riquelme, M., Jiménez-García, B., Abellán, N., Barba, R., Uribelarrea, D., Martín-Perea, D., Diez-Martin, F., Maíllo-Fernández, J. M., Rodríguez-Hidalgo, A., Courtenay, L., Mora, R., Maté-González, M. A., and González-Aguilera, D. 2021a: Early Pleistocene faunivorous hominins were not kleptoparasitic, and this impacted the evolution of human anatomy and socio-ecology. Scientific Reports 11:16135.
Domínguez-Rodrigo, M., Gidna, A., Baquedano, E., Cobo-Sánchez, L., Mora, R., Courtenay, L. A., Gonzalez-Aguilera, D., Mate-Gonzalez, M. A., and Prieto-Herráez, D. 2021b: A 3D taphonomic model of long bone modification by lions in medium-sized ungulate carcasses. Scientific Reports 11:4944.
Ferraro, J. V., Plummer, T. W., Pobiner, B. L., Oliver, J. S ., Bishop, L. C., Braun, D. R., Ditchfield, P. W., Seaman, J. W., III, Binetti, K. M., Seaman, J. W., Jr, Hertel, F., and Potts, R. 2013: Earliest Archaeological Evidence of Persistent Hominin Carnivory. PLoS ONE 8(4):e62174.
Foley, R. A. 1996: An Evolutionary and Chronological Framework for Human Social Behaviour. Proceedings of the British Academy 88, 95–117.
Galán, A. B., Rodríguez, M., de Juana, S., and Domínguez-Rodrigo, M. 2009: A new experimental study on percussion marks and notches and their bearing on the interpretation of hammerstone-broken faunal assemblages. Journal of Archaeological Science 36, 776–784.
Gaudzinski-Windheuser, S., Noack, E. S., Pop, E., Herbst, C., Pfleging, J., Buchli, J., Enzmann, F., Kindler, L., Iovita, R., Street, M., and Roebroeks, W. 2018: Evidence for close-range hunting by last interglacial Neanderthals. Nature Ecology & Evolution 2, 1087–1092.
Gidna, A., Domínguez-Rodrigo, M., and Pickering, T. R. 2015: Patterns of bovid long limb bone modification created by wild and captive leopards and their relevance to the elaboration of referential frameworks for paleoanthropology. Journal of Archaeological Science: Reports 2, 302–309.
Hay, R. L. 1976: Geology of the Olduvai Gorge: a study of sedimentation in a semiarid basin. London/Berkeley/Los Angeles: University of California Press.
Holliday, T. W. 2012: Body Size, Body Shape, and the Circumscription of the Genus Homo. Current Anthropology 53, Suppl. 6, S330–S345.
Isaac, G. 1978: The Food-sharing Behavior of Protohuman Hominids. Scientific American 238, 90–108.
Isaac, G. L. 1983: Aspects of human evolution. In: Isaac, B. (ed.), The archaeology of human origins. Papers by Glynn Isaac. Cambridge: Cambridge University Press, 96–119.
Jiménez-García, B., Aznarte, J., Abellán, N., Baquedano, E., and Domínguez-Rodrigo, M. 2020: Deep learning improves taphonomic resolution: high accuracy in differentiating tooth marks made by lions and jaguars. Journal of The Royal Society Interface 17: 20200446.
Leakey, M. D. 1971: Olduvai Gorge, Volume 3: Excavations in Beds I and II, 1960-1963. Cambridge: Cambridge University Press.
Lyman, R. L. 1994: Vertebrate Taphonomy. Cambridge: Cambridge University Press.
Manega, P. C. 1993: Geochronology, geochemistry and isotopic study of the Plio-Pleistocene hominid sites and the Ngorongoro Volcanic Highland in northern Tanzania. PhD dissertation, University of Colorado at Boulder.
Marlowe, F. W. 2010: The Hadza. Hunter-Gatherers of Tanzania. Berkeley: University of California Press.
Moclán, A. and Domínguez-Rodrigo, M. 2018: An experimental study of the patterned nature of anthropogenic bone breakage and its impact on bone surface modification frequencies. Journal of Archaeological Science 96, 1–13.
Moclán, A., Domínguez-Rodrigo, M., and Yravedra, J. 2019: Classifying agency in bone breakage: an experimental analysis of fracture planes to differentiate between hominin and carnivore dynamic and static loading using machine learning (ML) algorithms. Archaeological and Anthropological Sciences 11, 4663–4680.
Niiniluoto, I. 1987: Truthlikeness. Synthese Library 185. Dordrecht: D. Reidel Publishing Company.
Oliver, J. S., Plummer, T. W., Hertel, F., and Bishop, L. C. 2019: Bovid mortality patterns from Kanjera South, Homa Peninsula, Kenya and FLK-Zinj, Olduvai Gorge, Tanzania: Evidence for habitat mediated variability in Oldowan hominin hunting and scavenging behavior. Journal of Human Evolution 131, 61–75.
Organista, E., Moclán, A., Aramendi, J., Cobo-Sánchez, L., Egeland, C. P., Uribellarea, D., Martín-Perrea, D., Vegara-Riquelme, M., Hernández-Vivanco, L., Gidna, A., Mabulla, A., Baquedano, E., and Domínguez-Rodrigo, M. in press: A taphonomic analysis of PTK (Bed I, Olduvai Gorge) and its bearing on the interpretation of the dietary and ecospatial behaviors of early humans. Quaternary Science Reviews 300.
Parkinson, J. A. 2018: Revisiting the hunting-versus-scavenging debate at FLK Zinj: A GIS spatial analysis of bone surface modifications produced by hominins and carnivores in the FLK 22 assemblage, Olduvai Gorge, Tanzania. Palaeogeography, Palaeoclimatology, Palaeoecology 511, 29–51.
Parkinson, J. A., Plummer, T. W., Oliver, J. S., and Bishop, L. C. 2022: Meat on the menu: GIS spatial distribution analysis of bone surface damage indicates that Oldowan hominins at Kanjera South, Kenya had early access to carcasses. Quaternary Science Reviews 277: 107314.
Pickering, T. R. and Domínguez-Rodrigo, M. 2010: Chimpanzee Referents and the Emergence of Human Hunting. The Open Anthropology Journal 3, 107–113.
Plummer, T. W. and Bishop, L. C. 2016: Oldowan hominin behavior and ecology at Kanjera South, Kenya. Journal of Anthropological Sciences 94, 29–40.
Plummer, T. W., Bishop, L. C., Ditchfield, P. W., Ferraro, J. V., Kingston, J. D., Hertel, F., and Braun, D. R. 2009: The Environmental Context of Oldowan Hominin Activities at Kanjera South, Kenya. In: E. Hovers and D. R. Braun (eds.), Interdisciplinary Approaches to the Oldowan. Vertebrate Paleobiology and Paleoanthropology Series. Dordrecht: Springer, 149–160.
Pobiner, B. 2016: Meat-Eating Among the Earliest Humans. American Scientist 104, 110–117.
Pobiner, B. L., Rogers, M. J., Monahan, C. M., and Harris, J. W. K. 2008: New evidence for hominin carcass processing strategies at 1.5 Ma, Koobi Fora, Kenya. Journal of Human Evolution 55, 103–130.
Popper, K. 1972: The Logic of Scientific Discovery. Sixth (revised) impression. London: Hutchinson.
Rabinovich, R., Gaudzinski-Windheuser, S., and Goren-Inbar, N. 2008: Systematic butchering of fallow deer (Dama) at the early middle Pleistocene Acheulian site of Gesher Benot Ya’aqov (Israel). Journal of Human Evolution 54, 134–149.
Rodríguez-Hidalgo, A., Saladié, P., Ollé, A., and Carbonell, E. 2015: Hominin subsistence and site function of TD10.1 bone bed level at Gran Dolina site (Atapuerca) during the late Acheulean. Journal of Quaternary Science 30, 679–701.
Rodríguez-Hidalgo, A., Saladié, P., Ollé, A., Arsuaga, J. L., Bermúdez de Castro, J. M., and Carbonell, E. 2017: Human predatory behavior and the social implications of communal hunting based on evidence from the TD10.2 bison bone bed at Gran Dolina (Atapuerca, Spain). Journal of Human Evolution 105, 89–122.
Rose, L. and Marshall, F. 1996: Meat Eating, Hominid Sociality, and Home Bases Revisited. Current Anthropology 37, 307–338.
Sahnouni, M., Pares, J., Duval, M., van der Made, J., Harichane, Z., Perez-Gonzalez, A., Abdessadok, S., Arnold, L., Caceres, I., Kandi, N., Chelli-Cheheb, R., Boulaghraif, K., Agusti, J., Saidani, N., and Mouhoubi, Y. 2018: Early Homo adaptive and behavioral patterns in North Africa: perspectives from Ain Hanech and Tighennif (formerly Ternifine) sites in northern Algeria. Abstract, 15th Congress of PanAfrican Archaeological Association for Prehistory and Related Studies, Rabat, september 10-14, 2018, 5–6.
Saladié, P., Huguet, R., Díez, C., Rodríguez-Hidalgo, A., Cáceres, I., Vallverdú, J., Rosell, J., Bermúdez de Castro, J. M., and Carbonell, E. 2011: Carcass transport decisions in Homo antecessor subsistence strategies. Journal of Human Evolution 61, 425–446.
Saladié, P., Rodríguez-Hidalgo, A., Huguet, R., Cáceres, I., Díez, C., Vallverdú, J., Canals, A., Soto, M., Santander, B., Bermúdez de Castro, J. M., Arsuaga, J. L., and Carbonell, E. 2014: The role of carnivores and their relationship to hominin settlements in the TD6-2 level from Gran Dolina (Sierra de Atapuerca, Spain). Quaternary Science Reviews 93, 47–66.
Stancampiano, L. M., Sistiaga, A., Uribelarrea del Val, D., Aramendi, J., Baquedano, E., Mabulla, A., Domínguez-Rodrigo, M., and Magill, C. R. 2022: New site at Olduvai Gorge (AGS, Bed I, 1.84 Mya) widens the range of locations where hominins engaged in butchery. Scientific Reports 12:9794.
Stiner, M. C. 1990: The use of mortality patterns in archaeological studies of hominid predatory adaptations. Journal of Anthropological Archaeology 9, 305–351.
Treves, A. and Naughton-Treves, L. 1999: Risk and opportunity for humans coexisting with large carnivores. Journal of Human Evolution 36, 275–282.
Uribelarrea, D., Domínguez-Rodrigo, M., Pérez-González, A., Vegas Salamanca, J., Baquedano, E., Mabulla, A., Musiba, C., Barboni, D., and Cobo-Sánchez, L. 2014: Geo-archaeological and geometrically corrected reconstruction of the 1.84 Ma FLK Zinj paleolandscape at Olduvai Gorge, Tanzania. Quaternary International 322-323, 7–31.
Villmoare, B., Kimbel, W. H., Seyoum, C., Campisano, C. J., DiMaggio, E. N., Rowan, J., Braun, D. R., Arrowsmith, J. R., and Reed, K. E. 2015: Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. Science 347, 1352–1355.
Walter, R. C., Manega, P. C., Hay, R. L., Drake, R. E., and Curtis, G. H. 1991: Laser-fusion 40Ar/39Ar dating of Bed I, Olduvai Gorge, Tanzania. Nature 354,145–149.
Walter, R. C., Manega, P. C., and Hay, R. L. 1992: Tephrochronology of Bed I, Olduvai Gorge: An application of laserfusion 40Ar/39Ar dating to calibrating biological and climatic change. Quaternary International 13/14, 37–46.