Abstract
In the introductory chapter of this edited volume, I argue that paleoanthropological research on hominin behavioral evolution tends to overlook the conceptual distinction between a species’ basic anatomical capacity to carry out a certain physical task (e.g., the ability to climb), its evolved biomechanical efficiency in performing that activity (e.g., arboreal climbing efficiency), and each individual’s habitual physical activities (e.g., frequency and intensity of climbing throughout life). Using a few key examples from the literature, I posit that the lack of this theoretical distinction can compromise the integrity of paleoanthropological hypotheses and interpretations surrounding hominin biocultural evolution. Lastly, this chapter encourages future evolutionary studies to always strive to address all three behavioral components (capacity, evolved efficiency, and habitual behavior), relying on appropriate methods (and morphological traits) for each of them.
Introduction
Reconstructing habitual behavior in the past comprises a major objective of archaeological sciences centered on bio-cultural evolution. One of the most fundamental components of a population’s behavior involves its daily bodily activities, which form the physical expression of its cultural and subsistence practices in response to the dynamic conditions of the surrounding environment. Essentially, reconstructions of physical activity attempt to answer the fundamental question of “What did people do in the past?”, seeking to piece together mild and fragmentary reflections of hominin daily life, subsistence strategies, social hierarchy, symbolic behavior, and cultural practices. The importance of this difficult objective, which is sometimes referred to as the “Holy Grail of Bioarchaeology” (Jurmain et al. 2011), has led to the emergence of an entire field of evolutionary research, involving a plethora of proposed theoretical concepts, methods, and techniques (e.g., Karakostis and Harvati 2021; Kivell 2016; Wallace et al. 2017). Naturally, reconstructing human behavior based on dry skeletal remains is always an arduous endeavor due to the inherent complexity and diversity of human behavior, combined with the multifactorial etiology of bone morphological variation (Pearson and Lieberman 2004; Ruff et al. 2006; Schrader 2019; Wallace et al. 2017).
In evolutionary anthropology, numerous previous studies have relied on reconstructions of physical activity to address some of the most fundamental questions on human evolution, such as the emergence of obligate bipedal locomotion in early hominins (e.g., Daver et al. 2022; Richmond and Jungers 2008), the earliest evidence of habitual stone tool use (e.g., Karakostis et al. 2021; Kivell 2015; Marzke 1997, 2013), behavioral and cultural differences across recent hominins (e.g., Neanderthals and modern humans; see Bardo et al. 2020; Karakostis et al. 2018; Maki and Trinkaus 2011; Niewoehner 2006; Pearson et al. 2006), the tool-making abilities of enigmatic fossil hominins (i.e., Homo naledi, Homo floresiensis, and Homo luzonensis; see Détroit et al. 2019; Kivell et al. 2015; Tocheri et al. 2007), or the proposed emergence of key aspects of behavioral modernity in Homo sapiens, including division of labor, greater environmental adaptability, and the production of sophisticated artifacts (e.g., see Estalrrich and Rosas 2015; Karakostis et al. 2020; Niewoehner 2001).
Typically, most anthropological studies addressing these major evolutionary questions have relied on comparing skeletal functional morphology across diverse hominin species and comparative samples of extant species (e.g., Dunmore et al. 2020; Kivell 2015; Marzke 2013; Marzke et al. 2010). Such morphological markers of activity are associated either with species-wide evolutionary adaptations to increased biomechanical efficiency (e.g., bone shape and joint configurations that allow efficient climbing locomotion or dexterous “in-hand” manipulation of objects) or lifetime alterations in bone morphology due to phenotypic plasticity and biomechanical loading history (Ruff et al. 2006). The latter mainly involve variation in long bone cross-sectional geometry, trabecular morphology, pathological lesions potentially associated with intense physical stress (e.g., osteoarthritis), and the morphology of muscle attachment sites on the bone surface (or “entheses”) (e.g., Kivell 2016; Schrader 2019; Stock and Shaw 2007; Wallace et al. 2017). The basic premise of reconstructing activity based on most of these activity markers relies on the broad concept of “bone functional adaptation”, according to which bone form (size and shape) is expected to adapt to altered physical stimuli both before and after adulthood (Pearson and Lieberman 2004; Ruff et al. 2006).
Even though the reliability of many of these activity markers has been often questioned due to (mainly) their multifactorial etiology and the lack of supporting laboratory evidence (Schrader 2019; Wallace et al. 2017; Wallace et al. 2022), some of them have been repeatedly validated through extensive experimental work (e.g., Lieberman et al. 2004; Shaw and Stock 2009; Wallace et al. 2022). This includes a method I introduced in 2016 (Karakostis 2015; Karakostis and Lorenzo 2016), recently named the “Validated Entheses-based Reconstruction of Activity” (VERA) approach (see literature reviews by Karakostis 2022, and Karakostis and Harvati 2021), whose reliability has been supported in several studies involving diverse laboratory animals (e.g., Castro et al. 2022; Karakostis, Jeffery, et al. 2019, Karakostis, Wallace, et al. 2019; Karakostis and Wallace 2023) as well as human skeletons with a universally unique level of long-term occupational documentation (Karakostis et al. 2017; Karakostis and Hotz 2022). In this volume, a detailed critical review of most of the above activity markers has been provided in the chapter of this book authored by Dr. Jane Buikstra.
This chapter posits that the conceptual distinction between habitual physical activity and evolutionary functional adaptation is often unclear in the paleoanthropological literature centered on hominin biocultural evolution. In contrast, the above skeletal markers of “habitual activity” and “biomechanical efficiency” are typically lumped together as vague indicators of human behavior. This misconception, however, implies that an individual’s daily living conditions and activities directly reflect its species’ evolutionary history, thus underestimating the crucial effects of environmental and/or cultural factors on an individual’s behavior. Here, I argue that this practice directly compromises the reliability of our hypotheses and interpretations surrounding the evolution of hominin behavior. This commentary paper, which forms the introductory chapter of this edited volume, is divided into three main sections:
- Habitual activity versus evolved efficiency
- Fake it until you make it? Habitual tool use versus tool-using dexterity in early hominins
- Neanderthals: Strong yet precise?
Habitual activity versus evolved efficiency
By inferring hominin daily physical behavior based on skeletal markers of species-wide biomechanical efficiency (e.g., manual dexterity), it is effectively implied that hominin individuals habitually performed what they were best adapted for (from birth). This viewpoint blatantly overlooks the existence of ecological constraints that regulate the conditions of a population’s fitness within an everchanging environment, which are bound to vary greatly in time and space throughout a species’ evolutionary history. Moreover, in an evolutionary context, it seems unlikely that the efficient performance of an activity can be naturally selected before this behavior is even practiced at all (at least to some degree) by a species or population that had not yet developed such efficiency (with the exception of behaviors that depend on functional traits resulting from exaptation). Furthermore, using evolved traits to reconstruct habitual activity also underestimates the weight of cultural variability that may often be unrelated to the biomechanical constraints of our evolutionary history. This is especially the case for the more recent and larger-brained hominins (e.g., Neanderthals and early modern humans), who have been associated with sophisticated cultural practices (Villa and Roebroeks 2014).
The importance of conceptually distinguishing between skeletal markers of habitual activity (associated with phenotypic plasticity) and biomechanical efficiency (resulting from evolutionary adaptation) is graphically demonstrated in the example of Figure 1 (more information provided in its legend), which focuses on arboreal climbing behaviors in chimpanzees and modern humans (Fig. 1). As presented in that graph, humans and chimpanzees share the basic functional capacity for performing arboreal climbing (represented by the left circle of the figure). Nevertheless, due to a series of evolutionary species-wide adaptations, chimpanzee bodies are much more efficient in climbing trees (the highlighted area of the left circle). If this comparative framework was used to infer daily behavior in individuals of these species, the conclusion would be that no modern human is habitually involved in arboreal climbing. However, as shown in the examples of the two bottom pictures, there are humans who habitually climbed throughout their lives (and cases of chimpanzees that never did; see Wallace et al. 2020). The behaviors depicted in these examples are in line with the specific individuals’ environmental contexts and demands (represented by the right circle of the figure).

Evidently, these daily physical practices are not expected to silence the presence of genetically determined anatomical traits associated with climbing efficiency in a species (e.g., the occurrence of curved phalanges in chimpanzees), as previously demonstrated (e.g., Wallace et al. 2020). These habitual activities, however, might influence the morphology of bone traits affected by biomechanical stress throughout life (see section above), including internal bone structures and entheseal 3D surface morphology (e.g., Karakostis et al. 2019; Karakostis and Harvati 2021; Kivell 2016; Macintosh et al. 2017; Shaw and Stock 2009; Wallace et al. 2017).
“Fake it until you make it?”
: Habitual tool use versus tool-using dexterity in early hominins
The origins of human-like stone tool use represent one of the most crucial research questions in human evolutionary sciences. Traditionally, habitual stone tool use had been broadly associated with the emergence of the genus Homo. However, this viewpoint has now shifted due to the recent discovery of proposed stone tool industries predating our genus (Harmand et al. 2015) and the identification of australopithecine hand skeletons bearing anatomical indications of increased manual dexterity (e.g., Dunmore et al. 2020; Kivell 2015; Kivell et al. 2018). For instance, Australopithecus sediba and—to a lesser degree—Australopithecus afarensis exhibit a thumb that is proportionally much longer than in chimpanzees, arguably facilitating the efficient performance of human-like precision grips involving interactions between the thumb and the fingers (Kivell et al. 2018). Relying on such observations, previous research suggested that these species may have been the producers and users of early stone tool industries (Kivell 2015). Based on a similar theoretical principle, a previous biomechanical modeling study proposed that Australopithecus afarensis was likely not the producer of the oldest proposed stone tools (the Lomekwian industry) because of its fifth digit’s low force-producing efficiency, which might have prevented it from habitually manipulating the large Lomekwian cores and tools (Domalain et al. 2017).
However, despite the great importance of these functional adaptations and their crucial implications for the biomechanical evolution of hominin dexterity, the etiology of these traits seems to be unrelated to phenotypic plasticity and daily manual behavior. As summarized in Figure 2, all hominins seem to have been capable of human-like precision grasping, regardless of the suggested comparatively low efficiency of some hominins (e.g., Australopithecus) in opposing their thumb and fifth rays (Domalain et al. 2017; Hopkins et al. 2002; Karakostis et al. 2021; Marzke et al. 1999). Assuming that the cognitive capacity of a fossil hominin species would have permitted its individuals to mentally conceive the production of stone tools, and that using tools would have been both feasible and beneficial within a particular environmental context, it seems nonsensical that these individuals would decide not to use stone tools because their hands were comparatively less dexterous than those of another hominin species (i.e., our comparative samples in an analysis). In fact, considering that early hominins were probably fully capable of precise manipulation (such as all extant great apes; Hopkins et al. 2002), it seems very unlikely that they would only start using stone tools after their manual efficiency gradually increased. In contrast, unless hominin tool-using behaviors arose and evolved exclusively through exaptation, it seems more plausible that the adaptive value of tool-using dexterity grew in hominins who were already using stone tools (at least to some degree) and whose fitness could thus directly benefit from higher manipulatory efficiency (see discussion in Kunze et al. 2022).
In two recent studies (Karakostis et al. 2021; Kunze et al. 2022), my collaborators and I addressed early hominin biomechanical efficiency and habitual behavior separately, using different methods for each of the two behavioral components. In Karakostis et al. (2021), we relied on an integrative biomechanical modeling approach for directly calculating thumb opposition efficiency considering the effects of both the missing muscle architecture and bone morphology. The results showed that the earliest proposed tool-using species, including Australopithecus sediba, showed a relatively low thumb opposition dexterity. In fact, when our models assumed a chimpanzee-like muscle force-producing capacity, the thumb dexterity of that species was similar to that of extant chimpanzees. Nevertheless, the application of the method VERA on the hand entheses of Australopithecus sediba indicated the habitual use of a muscle that is essential for human-like stone tool use (Kunze et al. 2022), while another study showed that its thumb’s trabecular morphology was found to be consistent with human-like manipulation (Dunmore et al. 2020). Altogether, these findings suggest that the individual representing this hominin species may have frequently performed human-like hand grips regardless of its comparatively low thumb dexterity (Kunze et al. 2022; also see plot at the bottom of Fig. 2). These opposing trends between reconstructed efficiency and habitual behavior in Australopithecus sediba highlight the importance of distinguishing between these two notions (and associated analytical methods) when approaching the evolution of hominin behavior.

Neanderthals: strong yet precise?
Another interesting example relates to the manual behaviors of Neanderthals. In comparison to modern humans, Neanderthal hand bones tend to be more robust and bear more pronounced muscle attachment sites (Niewoehner 2006). Moreover, their thumb presented a relatively (slightly) shorter length, different proportions between the two adjoining phalanges, and a basal (trapeziometacarpal) joint configuration consistent with less pollical flexibility in some respects (Churchill 2001; Niewoehner 2001, 2006; Niewoehner et al. 2003). Based on these anatomical indications, previous studies hypothesized that the Neanderthal lifestyle predominantly relied on transverse power-grasping motions, which were in turn associated with the use of composite tools and hafting (Niewoehner 2006). More recently, this hypothesis of habitual use of hafted tools received additional support from a biomechanical study focusing on the thumb’s basal joint (Bardo et al. 2020), which reported that Neanderthal thumbs were better adapted for performing thumb extension (i.e., a central component of power grasping) compared to other hand movements (e.g., the opposition of the thumb for human-like precision grasping).
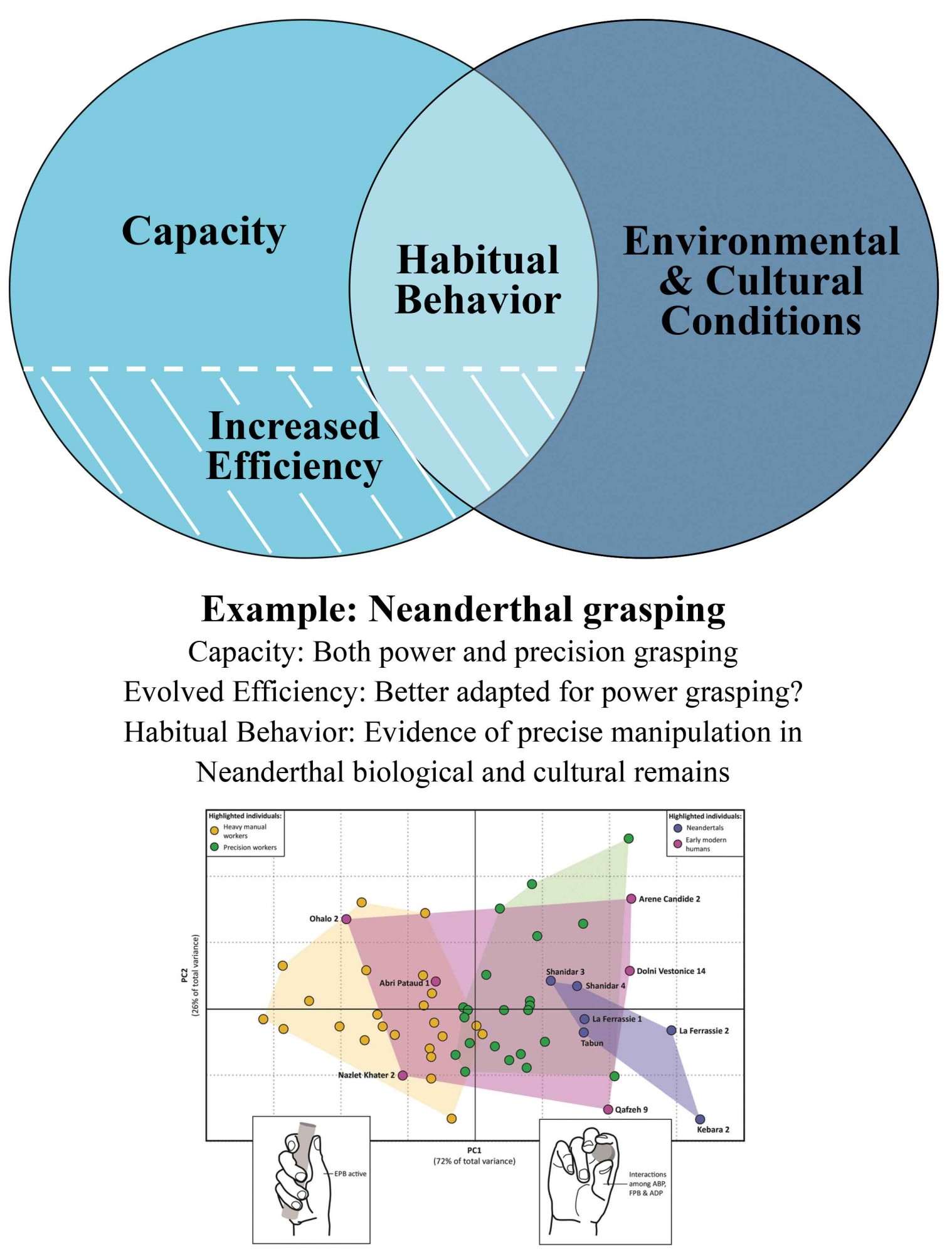
However, it could be argued that the above anatomical configurations are of genetic origin, occurring across Neanderthal skeletons from extremely diverse geochronological and environmental contexts (Niewoehner 2006). Furthermore, previous biomechanical modeling studies demonstrated that Neanderthal hands were perfectly capable of performing modern human-like precision grasping involving small objects (Feix et al. 2015; Karakostis et al. 2021; Niewoehner et al. 2003). On this basis, by hypothesizing that the above anatomical traits can be used to reconstruct daily manual activity in Neanderthals, it is indirectly—yet clearly—assumed that their presumed power-grasping adaptations would lead them to systematically prefer power grips even in environmental contexts necessitating precision grasping (for which they were anatomically capable). This hypothesis contradicts experimental studies suggesting that producing and manipulating the vast majority of small stone tools commonly associated with Neanderthal contexts (e.g., Mousterian flakes) would primarily require the use of thumb-index precision grasping (Key and Lycett 2018). In contrast, direct evidence of hafting (presumably associated with transverse power grasping) remains extremely scarce across Neanderthal contexts, while being entirely absent in many Neanderthal sites with rich lithic assemblages (Claud et al. 2019; Niewoehner 2006). Finally, there is an increasing body of archaeological evidence suggesting that Neanderthals were engaging in various practices requiring a high level of manual precision, such as the production of specialized bone tools, making of cordage, and the use of tar (Hardy et al. 2013; Soressi et al. 2013; Villa and Roebroeks 2014).
In a previous study (Karakostis et al. 2018), my collaborators and I sought to address this controversy between the above-mentioned findings of biomechanical modeling (showing that Neanderthals had the capacity to precisely manipulate small objects), archaeological evidence (involving the use of predominantly “microlithic” tools and precise manual practices), and functional indications of efficiency (showing that Neanderthal hands may have been better adapted for transverse power grasping). This was attempted through the application of my experimentally validated VERA method (Karakostis 2015, 2022) on a diverse comparative sample of Neanderthals, early modern humans, and recent individuals with extensively documented life histories. The results showed clear evidence of habitual thumb-index precision grasping in Neanderthals, contradicting the traditional viewpoint that Neanderthals habitually relied on power grasping (reflected on their high bone robusticity), and reflecting the latest archaeological indications on the daily manual behavior of this species. Figure 3 summarizes the proposed distinction among Neanderthal capacity, efficiency (dexterity), and habitual behavior for precision grasping behaviors.
Concluding remarks
The examples provided in this chapter posit that anthropological hypotheses and interpretations surrounding fossil hominin behavior could greatly benefit from properly distinguishing among the fundamental concepts of broader anatomical capacity, evolved biomechanical efficiency, and habitual physical activity (Figs. 1 to 3). This distinction can be facilitated by the selection of appropriate methods for addressing each of these three basic components of physical behavior. Anatomical capacity and efficiency can be best evaluated based on biomechanical modeling techniques, which can be used to directly assess the functional significance of genetically determined morphological traits (e.g., species-wide overall bone robusticity trends, joint configuration/orientation, or consistent proportions among different bone elements), while also attempting to account (as much as possible) for the potential influence of the missing soft tissue (e.g., Karakostis et al. 2021). However, reconstructing habitual physical activities of an individual based on its species-wide evolved functional traits is highly misleading, as it blatantly overlooks the dynamic environmental and/or cultural factors affecting daily hominin behavior and subsistence strategies within each species. In contrast, that objective can be more reliably addressed by studying bone structures that are experimentally shown to reflect biomechanical loading history throughout life (for a more detailed perspective, see this book’s dedicated chapters by Jane Buikstra and Ian Wallace). Any resulting indication on habitual physical activity should always be interpreted on the basis of each species’ possible constraints (anatomical and cognitive capacity) and evolved biomechanical efficiency (e.g., level of dexterity), focusing on adequately reconstructed paleoenvironmental and/or cultural contexts (Fig. 1).
Acknowledgments
First, I am deeply thankful to all participants of the Symposium “Biocultural Implications: An Agenda for Integrative Approaches” (which took place in 2021 at the DFG Centre for Advanced Studies “Words, Bones, Genes, Tools”), and especially those contributing to the present edited volume. I am also thankful to all my collaborators worldwide, who made the research discussed in this review possible. Finally, special thanks are due to Miri Mertner for her substantial contribution in co-organizing the Symposium.
References
Bardo, A., M. H. Moncel, C. J. Dunmore, T. L. Kivell, E. Pouydebat, and R. Cornette. 2020. The Implications of Thumb Movements for Neanderthal and Modern Human Manipulation. Scientific Reports 10 (1): 19323. doi:10.1038/s41598-020-75694-2.
Castro, A. A., F. A. Karakostis, L. E. Copes, H. E. McClendon, A. P. Trivedi, N. E. Schwartz, and T. Garland Jr. 2022. Effects of Selective Breeding for Voluntary Exercise, Chronic Exercise, and Their Interaction on Muscle Attachment Site Morphology in House Mice. Journal of Anatomy 240 (2): 279–295. doi:10.1111/joa.13547.
Churchill, S. E. 2001. Hand Morphology, Manipulation, and Tool Use in Neandertals and Early Modern Humans of the Near East. Proceedings of the National Academy of Sciences 98 (6): 2953–2955. doi:10.1073/pnas.061032198.
Claud, É., C. Thiébaut, S. Costamagno, M. Deschamps, M. C. Soulier, M. Brenet, M. G. Chacón-Navarro, D. Colonge, A. Coudenneau, C. Lemorini, et al. 2019. The Practices Used by the Neanderthals in the Aquisition and Exploitation of Plant and Animal Resources and the Function of the Sites Studied: Summary and Discussion. Palethnologie. Archéologie et Sciences Humaines, no. 10 (November). Presses universitaires du Midi. doi:10.4000/palethnologie.4179.
Daver, G., F. Guy, H. T. Mackaye, A. Likius, J.-R. Boisserie, A. Moussa, L. Pallas, P. Vignaud, and N. D. Clarisse. 2022. Postcranial Evidence of Late Miocene Hominin Bipedalism in Chad. Nature 609 (7925): 94–100. doi:10.1038/s41586-022-04901-z.
Détroit, F., A. S. Mijares, J. Corny, G. Daver, C. Zanolli, E. Dizon, E. Robles, R. Grün, and P. J. Piper. 2019. A New Species of Homo from the Late Pleistocene of the Philippines. Nature 568 (7751): 181–186. doi:10.1038/s41586-019-1067-9.
Domalain, M., A. Bertin, and G. Daver. 2017. Was Australopithecus Afarensis Able to Make the Lomekwian Stone Tools? Towards a Realistic Biomechanical Simulation of Hand Force Capability in Fossil Hominins and New Insights on the Role of the Fifth Digit. Comptes Rendus Palevol, Hominin biomechanics, virtual anatomy and inner structural morphology: From head to toe. A tribute to Laurent Puymerail, 16 (5): 572–584. doi:10.1016/j.crpv.2016.09.003.
Dunmore, C. J., M. M. Skinner, A. Bardo, L. R. Berger, J. J. Hublin, D. H. Pahr, A. Rosas, N. B. Stephens, and T. L. Kivell. 2020. The Position of Australopithecus Sediba within Fossil Hominin Hand Use Diversity. Nature Ecology & Evolution 4 (7): 911–918. doi:10.1038/s41559-020-1207-5.
Estalrrich, A., and A. Rosas. 2015. Division of Labor by Sex and Age in Neandertals: An Approach through the Study of Activity-Related Dental Wear. Journal of Human Evolution 80: 51–63. doi:10.1016/j.jhevol.2014.07.007.
Feix, T., T. L. Kivell, E. Pouydebat, and A. M. Dollar. 2015. Estimating Thumb–Index Finger Precision Grip and Manipulation Potential in Extant and Fossil Primates. Journal of The Royal Society Interface 12 (106): 20150176. doi:10.1098/rsif.2015.0176.
Hardy, B. L., M. H. Moncel, C. Daujeard, P. Fernandes, P. Béarez, E. Desclaux, M. G. Chacon Navarro, S. Puaud, and R. Gallotti. 2013. Impossible Neanderthals? Making String, Throwing Projectiles and Catching Small Game during Marine Isotope Stage 4 (Abri Du Maras, France). Quaternary Science Reviews 82: 23–40. doi:10.1016/j.quascirev.2013.09.028.
Harmand, S., J. E. Lewis, C. S. Feibel, C. J. Lepre, S. Prat, A. Lenoble, X. Boës, R. L. Quinn, M. Brenet, A. Arroyo, et al. 2015. 3.3-Million-Year-Old Stone Tools from Lomekwi 3, West Turkana, Kenya. Nature 521 (7552): 310–315. doi:10.1038/nature14464.
Hopkins, W. D., C. Cantalupo, M. J. Wesley, A. B. Hostetter, and D. L. Pilcher. 2002. Grip Morphology and Hand Use in Chimpanzees (Pan Troglodytes). Journal of Experimental Psychology 131 (3): 412–423. doi:10.1037//0096-3445.131.3.412.
Jurmain, R., F. A. Cardoso, C. Henderson, and S. Villotte. 2011. Bioarchaeology’s Holy Grail: The Reconstruction of Activity. In A Companion to Paleopathology, 531–552. John Wiley & Sons, Ltd. doi:10.1002/9781444345940.ch29.
Karakostis, F. A. 2015. Morphometric Analysis of Entheseal Surfaces in Metacarpals, Proximal Hand Phalanges and Pollical Distal Phalanges from Modern Human Populations. Master Thesis, Tarragona, Spain: University of Rovira i Virgili.
Karakostis, F. A. 2022. Statistical Protocol for Analyzing 3D Muscle Attachment Sites Based on the “Validated Entheses based Reconstruction of Activity” (VERA) Approach. International Journal of Osteoarchaeology. doi: 10.1002/oa.3196.
Karakostis, F. A., D. Haeufle, I. Anastopoulou, K. Moraitis, G. Hotz, V. Tourloukis, and K. Harvati. 2021. Biomechanics of the Human Thumb and the Evolution of Dexterity. Current Biology 31: 1317–1325. doi:10.1016/j.cub.2020.12.041.
Karakostis, F. A., and K. Harvati. 2021. New Horizons in Reconstructing Past Human Behavior: Introducing the “Tübingen University Validated Entheses-Based Reconstruction of Activity” Method. Evolutionary Anthropology 30: 185–198. doi: 10.1002/evan.21892.
Karakostis, F. A., and G. Hotz. 2022. Reflections of Manual Labor in the Hand Entheses of Early Industrial Women Workers with Extensively Documented Life Histories. American Journal of Biological Anthropology. doi:10.1002/ajpa.24636.
Karakostis, F. A., G. Hotz, H. Scherf, J. Wahl, and K. Harvati. 2017. Occupational Manual Activity Is Reflected on the Patterns among Hand Entheses. American Journal of Physical Anthropology 164 (1): 30–40. doi:10.1002/ajpa.23253.
Karakostis, F. A., G. Hotz, V. Tourloukis, and K. Harvati. 2018. Evidence for Precision Grasping in Neandertal Daily Activities. Science Advances 4 (9): eaat2369. doi:10.1126/sciadv.aat2369.
Karakostis, F. A., N. Jeffery, and K. Harvati. 2019. Experimental Proof That Multivariate Patterns among Muscle Attachments (Entheses) Can Reflect Repetitive Muscle Use. Sci. Rep. 9 (1): 1–9. doi:10.1038/s41598-019-53021-8.
Karakostis, F. A., and C. Lorenzo. 2016. Morphometric Patterns among the 3D Surface Areas of Human Hand Entheses. American Journal of Physical Anthropology 160 (4): 694–707. doi:10.1002/ajpa.22999.
Karakostis, F. A., H. Reyes-Centeno, M. Francken, G. Hotz, K. Rademaker, and K. Harvati. 2020. Biocultural Evidence of Precise Manual Activities in an Early Holocene Individual of the High-Altitude Peruvian Andes. American Journal of Physical Anthropology 174: 35–48. doi:10.1002/ajpa.24160.
Karakostis, F. A., and I. J. Wallace. 2023. Climbing influences entheseal morphology in the humerus of mice: An experimental application of the VERA methodology. American Journal of Biological Anthropology 181 (1): 130–139. doi: 10.1002/ajpa.24700.
Karakostis, F. A., I. J. Wallace, N. Konow, and K. Harvati. 2019. Experimental Evidence That Physical Activity Affects the Multivariate Associations among Muscle Attachments (Entheses). Journal of Experimental Biology 222 (23): 213058. doi:10.1242/jeb.213058.
Key, A. J.M., and S. J. Lycett. 2018. Investigating Interrelationships between Lower Palaeolithic Stone Tool Effectiveness and Tool User Biometric Variation: Implications for Technological and Evolutionary Changes. Archaeological and Anthropological Sciences 10 (5): 989–1006. doi:10.1007/s12520-016-0433-x.
Kivell, T. L. 2015. Evidence in Hand: Recent Discoveries and the Early Evolution of Human Manual Manipulation. Philosophical Transactions of the Royal Society B 370 (1682): 20150105. doi:10.1098/rstb.2015.0105.
Kivell, T. L. 2016. A Review of Trabecular Bone Functional Adaptation: What Have We Learned from Trabecular Analyses in Extant Hominoids and What Can We Apply to Fossils? Journal of Anatomy 228 (4): 569–594. doi:10.1111/joa.12446.
Kivell, T. L., S. E. Churchill, J. M. Kibii, P. Schmid, and L. R. Berger. 2018. The Hand of Australopithecus Sediba. PaleoAnthropology 2018: 282–333.
Kivell, T. L., A. S. Deane, M. W. Tocheri, C. M. Orr, P. Schmid, J. Hawks, L. R. Berger, and S. E. Churchill. 2015. The Hand of Homo Naledi. Nature Communications 6 (1): 1–9. doi:10.1038/ncomms9431.
Kunze, J., F. A. Karakostis, S. Merker, M. Peresani, V. Tourloukis, and K. Harvati. 2022. Evidence for Habitual Tool Use in Early Hominins. PaleoAnthropology 2022 (2): 195–210. doi: 10.48738/2022.iss2.61.
Lieberman, D. E., J. D. Polk, and B. Demes. 2004. Predicting Long Bone Loading from Cross-Sectional Geometry. American Journal of Physical Anthropology 123 (2): 156–171. doi:10.1002/ajpa.10316.
Macintosh, A. A., R. Pinhasi, and J. T. Stock. 2017. Prehistoric Women’s Manual Labor Exceeded That of Athletes through the First 5500 Years of Farming in Central Europe. Science Advances 3 (11): eaao3893. doi:10.1126/sciadv.aao3893.
Maki, J., and E. Trinkaus. 2011. Opponens Pollicis Mechanical Effectiveness in Neandertals and Early Modern Humans. PaleoAnthropology 2011: 62–71.
Marzke, M. W. 1997. Precision Grips, Hand Morphology, and Tools. American Journal of Physical Anthropology 102 (1): 91–110.
Marzke, M. W. 2013. Tool Making, Hand Morphology and Fossil Hominins. Philosophical Transactions of the Royal Society B 368 (1630): 20120414. doi:10.1098/rstb.2012.0414.
Marzke, M. W., R. F. Marzke, R. L. Linscheid, P. Smutz, B. Steinberg, S. Reece, and K. N. An. 1999. Chimpanzee Thumb Muscle Cross Sections, Moment Arms and Potential Torques, and Comparisons with Humans. American Journal of Physical Anthropology 110 (2): 163–178. doi:10.1002/(SICI)1096-8644(199910).
Marzke, M. W., M. W. Tocheri, B. Steinberg, J. D. Femiani, S. P. Reece, R. L. Linscheid, C. M. Orr, and R. F. Marzke. 2010. Comparative 3D Quantitative Analyses of Trapeziometacarpal Joint Surface Curvatures among Living Catarrhines and Fossil Hominins. American Journal of Physical Anthropology 141 (1): 38–51. doi:10.1002/ajpa.21112.
Niewoehner, W. A. 2001. Behavioral Inferences from the Skhul/Qafzeh Early Modern Human Hand Remains. Proceedings of the National Academy of Sciences of the United States of America 98: 2979–1984. doi:10.1073/pnas.041588898.
Niewoehner, W. A. 2006. Neanderthal Hands in Their Proper Perspective. In Neanderthals Revisited: New Approaches and Perspectives, ed. by J.J. Hublin, K. Harvati, and T. Harrison, pp. 157–190. Vertebrate Paleobiology and Paleoanthropology. Dordrecht: Springer Netherlands. doi:10.1007/978-1-4020-5121-0_9.
Niewoehner, W. A., A. Bergstrom, D. Eichele, M. Zuroff, and J. T. Clark. 2003. Manual Dexterity in Neanderthals. Nature 422 (6930): 395–395. doi:10.1038/422395a.
Pearson, O. M., R. M. Cordero, and A. M. Busby. 2006. How Different Were Neanderthals’ Habitual Activities? A Comparative Analysis with Diverse Groups of Recent Humans. In Neanderthals Revisited: New Approaches and Perspectives, ed. by J.-J. Hublin, K. Harvati, and T. Harrison, pp. 135–156. Vertebrate Paleobiology and Paleoanthropology. Dordrecht: Springer Netherlands. doi:10.1007/978-1-4020-5121-0_8.
Pearson, O. M., and D. E. Lieberman. 2004. The Aging of Wolff’s “Law”: Ontogeny and Responses to Mechanical Loading in Cortical Bone. American Journal of Physical Anthropology Suppl 39: 63–99. doi:10.1002/ajpa.20155.
Richmond, B. G., and W. L. Jungers. 2008. Orrorin Tugenensis Femoral Morphology and the Evolution of Hominin Bipedalism. Science 319 (5870): 1662–1665.
doi:10.1126/science.1154197.
Ruff, C., B. Holt, and E. Trinkaus. 2006. Who’s Afraid of the Big Bad Wolff?: “Wolff’s Law” and Bone Functional Adaptation. American Journal of Physical Anthropology 129 (4): 484–498. doi:10.1002/ajpa.20371.
Schrader, S. 2019. Activity, Diet and Social Practice: Addressing Everyday Life in Human Skeletal Remains. Bioarchaeology and Social Theory. New York: Springer International Publishing. doi:10.1007/978-3-030-02544-1.
Shaw, C. N., and J. T. Stock. 2009. Habitual Throwing and Swimming Correspond with Upper Limb Diaphyseal Strength and Shape in Modern Human Athletes. American Journal of Physical Anthropology 140 (1): 160–172. doi:10.1002/ajpa.21063.
Soressi, M., S. P. McPherron, M. Lenoir, T. Dogandžić, P. Goldberg, Z. Jacobs, Y. Maigrot, N. L. Martisius, C. E. Miller, W. Rendu, et al. 2013. Neandertals Made the First Specialized Bone Tools in Europe. Proceedings of the National Academy of Sciences 110 (35): 14186–14190. doi:10.1073/pnas.1302730110.
Stock, J. T., and C. N. Shaw. 2007. Which Measures of Diaphyseal Robusticity Are Robust? A Comparison of External Methods of Quantifying the Strength of Long Bone Diaphyses to Cross-Sectional Geometric Properties. American Journal of Physical Anthropology 134 (3): 412–423. doi:10.1002/ajpa.20686.
Tocheri, M. W., C. M. Orr, S. G. Larson, T. Sutikna, J., E. Wahyu Saptomo, R. Awe Due, T. Djubiantono, M. J. Morwood, and W. L. Jungers. 2007. The Primitive Wrist of Homo Floresiensis and Its Implications for Hominin Evolution. Science 317 (5845): 1743–1745. doi:10.1126/science.1147143.
Villa, P., and W. Roebroeks. 2014. Neandertal Demise: An Archaeological Analysis of the Modern Human Superiority Complex. PLOS ONE 9 (4): e96424.
doi:10.1371/journal.pone.0096424.
Wallace, I. J., M. L. Burgess, and B. A. Patel. 2020. Phalangeal Curvature in a Chimpanzee Raised like a Human: Implications for Inferring Arboreality in Fossil Hominins. Proceedings of the National Academy of Sciences 117 (21): 11223–11225. doi:10.1073/pnas.2004371117.
Wallace, I. J., B. Demes, and S. Judex. 2017. Ontogenetic and Genetic Influences on Bone’s Responsiveness to Mechanical Signals. In Building Bones: Bone Formation and Development in Anthropology, ed. by C. J. Percival and J. T. Richtsmeier, pp. 234–253. Cambridge: Cambridge University Press.
Wallace, I. J., G. J. Riew, R. Landau, A. M. Bendele, N. B. Holowka, T. L. Hedrick, N. Konow, D. J. Brooks, and D. E. Lieberman. 2022. Experimental Evidence That Physical Activity Inhibits Osteoarthritis: Implications for Inferring Activity Patterns from Osteoarthritis in Archeological Human Skeletons. American Journal of Biological Anthropology 177 (2): 223–231. doi:10.1002/ajpa.24429.
Wallace, I. J., S. Worthington, D. T. Felson, R. D. Jurmain, K. T. Wren, H. Maijanen, R. J. Woods, and D. E. Lieberman. 2017. Knee Osteoarthritis Has Doubled in Prevalence since the Mid-20th Century. Proceedings of the National Academy of Sciences 114 (35): 9332–9336. doi:10.1073/pnas.1703856114.